Melanina
La melanina (del griego antiguo μέλας (mélas) ‘negro, oscuro’) es una familia de biomoléculas organizadas como oligómeros o polímeros, que entre otras funciones proporcionan los pigmentos de muchos organismos.[1] Los pigmentos de melanina se producen en un grupo especializado de células conocidas como melanocitos.


Existen cinco tipos básicos de melanina: eumelanina, feomelanina, neuromelanina, alomelanina y piomelanina.[2] La eumelanina se produce mediante un proceso químico de varias etapas conocido como melanogénesis, en el que a la oxidación del aminoácido tirosina le sigue la polimerización. La eumelanina es el tipo más común. La feomelanina, que se produce cuando los melanocitos funcionan mal debido a la derivación del gen a su formato recesivo, es un derivado de la cisteína que contiene porciones de polibenzotiazina que son responsables en gran medida del tinte rojo o amarillo que adquieren algunos colores de piel o cabello. La neuromelanina se encuentra en el cerebro. Se ha investigado su eficacia en el tratamiento de trastornos neurodegenerativos como el Parkinson.[3] La alomelanina y la piomelanina son dos tipos de melanina sin nitrógeno.
En la piel humana, la melanogénesis se inicia con la exposición a la radiación ultravioleta (UV), lo que provoca el oscurecimiento de la piel. La eumelanina es un eficaz absorbente de la luz; el pigmento es capaz de disipar más del 99,9% de la radiación UV absorbida.[4] Debido a esta propiedad, se cree que la eumelanina protege las células de la piel de los daños causados por la radiación UVA y UVB, reduciendo el riesgo de agotamiento de folato y degradación dérmica. La exposición a la radiación UV se asocia a un mayor riesgo de melanoma maligno, un cáncer de melanocitos (células de melanina). Los estudios han demostrado una menor incidencia de cáncer de piel en individuos con mayor concentración de melanina, es decir, con un tono de piel más oscuro.[5]
Tipos de melanina
editarEumelanina
editar
La eumelanina tiene dos formas ligadas al 5,6-dihidroxiindol (DHI) y al ácido 5,6-dihidroxiindol-2-carboxílico (DHICA). La eumelanina derivada del DHI es de color marrón oscuro o negro e insoluble, y la derivada del DHICA es más clara y soluble en álcali. Ambas eumelaninas surgen de la oxidación de la tirosina en orgánulos especializados llamados melanosomas. Esta reacción está catalizada por la enzima tirosinasa. El producto inicial, la dopaquinona, puede transformarse en 5,6-dihidroxiindol (DHI) o ácido 5,6-dihidroxiindol-2-carboxílico (DHICA). El DHI y el DHICA se oxidan y polimerizan para formar las dos eumelaninas.[6]
En condiciones naturales, la DHI y la DHICA a menudo copolimerizan, dando lugar a una gama de polímeros de eumelanina. Estos polímeros contribuyen a la variedad de componentes de la melanina en la piel y el pelo humanos, que van desde la feomelanina amarillo claro/rojo a la eumelanina enriquecida con DHICA marrón claro y la eumelanina enriquecida con DHI marrón oscuro o negro. Estos polímeros finales difieren en solubilidad y color.[6]
El análisis de la piel muy pigmentada (tipos V y VI en la escala de Fitzpatrick) revela que la eumelanina DHI constituye la mayor parte, aproximadamente un 60-70 %, seguida de la eumelanina DHICA, con un 25-35 %, y la feomelanina, sólo un 2-8 %. En particular, mientras que durante el bronceado se produce un enriquecimiento de la eumelanina DHI, éste va acompañado de una disminución de la eumelanina DHICA y la feomelanina.[6] Una pequeña cantidad de eumelanina negra en ausencia de otros pigmentos provoca la aparición de canas. Una pequeña cantidad de eumelanina en ausencia de otros pigmentos provoca el pelo rubio.[7] La eumelanina está presente en la piel, el pelo, etc.
Feomelanina
editar
Las feomelaninas (o feomelaninas) confieren una gama de colores que van del amarillento al rojizo.[8] Las feomelaninas se concentran especialmente en los labios, los pezones, el glande del pene y la vagina.[9] Cuando una pequeña cantidad de eumelanina en el pelo (que de otro modo daría lugar a un pelo rubio) se mezcla con la feomelanina, el resultado es un pelo anaranjado, que suele denominarse «pelirrojo». La feomelanina también está presente en la piel, por lo que los pelirrojos también suelen tener un tono más rosado. La exposición de la piel a la luz ultravioleta aumenta el contenido de feomelanina, al igual que ocurre con la eumelanina; pero en lugar de absorber la luz, la feomelanina del pelo y la piel refleja la luz entre amarilla y roja, lo que puede aumentar el daño causado por la exposición a la radiación UV.[10]
La producción de feomelanina depende en gran medida de la disponibilidad de cisteína, que se transporta al melanosoma y reacciona con la dopaquinona para formar cis-dopa. A continuación, la cis-dopa sufre varias transformaciones antes de formar la feomelanina.[6] En términos químicos, las feomelaninas difieren de las eumelaninas en que la estructura del oligómero incorpora unidades de benzotiazina y benzotiazol que se producen,[11] en lugar de DHI y DHICA, cuando está presente el aminoácido L-cisteína.
Neuromelanina
editarLa neuromelanina (NM) es un pigmento polimérico insoluble producido en poblaciones específicas de neuronas catecolaminérgicas del cerebro. Los humanos tienen la mayor cantidad de NM, que está presente en menor cantidad en otros primates y totalmente ausente en muchas otras especies.[12] La función biológica sigue siendo desconocida, aunque se ha demostrado que la NM humana se une eficazmente a metales de transición como el hierro, así como a otras moléculas potencialmente tóxicas. Por lo tanto, puede desempeñar un papel crucial en la apoptosis y la enfermedad de Parkinson relacionada.[13]
Otras formas de melaninas
editarHasta la década de 1960, la melanina se clasificaba en eumelanina y feomelanina. Sin embargo, en 1955 se descubrió una melanina asociada a las células nerviosas, la neuromelanina. En 1972 se descubrió una forma hidrosoluble, la piomelanina. En 1976 se encontró en la naturaleza la alomelanina, la quinta forma de las melaninas.[2]
Péptidomelanina
editar

La péptidomelanina es otra forma hidrosoluble de melanina,[14] que las esporas germinadas de Aspergillus niger (cepa: melanoliber) secretan en el medio circundante. La péptidomelanina se forma como un copolímero entre la L-DOPA eumelanina y péptidos cortos que forman una «corona», responsables de la solubilidad de la sustancia. Las cadenas peptídicas están unidas al polímero central de L-DOPA mediante enlaces peptídicos. Esto conduce a un proceso biosintético propuesto que implica la hidroxilación de péptidos tirosinilados formados a través de proteasas durante la esporogénesis, que luego se incorporan autoxidativamente a un polímero central de L-DOPA en crecimiento.
Selenomelanina
editarEs posible enriquecer la melanina con selenio en lugar de azufre. Este análogo de selenio de la feomelanina se ha sintetizado con éxito mediante rutas químicas y biosintéticas utilizando selenocistina como materia prima.[15] Debido al mayor número atómico del selenio, cabe esperar que la selenomelanina obtenida proporcione una mejor protección contra la radiación ionizante en comparación con las otras formas conocidas de melanina. Esta protección se ha demostrado con experimentos de radiación en células humanas y bacterias, lo que abre la posibilidad de aplicaciones en viajes espaciales.[16]
Tricocromos
editarLos tricocromos (antes llamados tricosiderinas) son pigmentos producidos a partir de la misma vía metabólica que las eumelaninas y las feomelaninas, pero a diferencia de estas moléculas tienen bajo peso molecular. Se encuentran en algunos cabellos rojos humanos.[17]
Humanos
editar
En los seres humanos, la melanina es el principal determinante del color de la piel. También se encuentra en el pelo, el tejido pigmentado subyacente al iris del ojo y la estría vascular del oído interno. En el cerebro, los tejidos que contienen melanina son la médula y las neuronas pigmentadas del tronco encefálico, como el locus cerúleo. También se encuentra en la zona reticular de la glándula suprarrenal.[18]
La melanina de la piel es producida por los melanocitos, que se encuentran en el estrato basal de la epidermis. Aunque, en general, los seres humanos poseen una concentración similar de melanocitos en la piel, los melanocitos de algunos individuos y grupos étnicos producen cantidades variables de melanina. La proporción de eumelanina (74 %) y feomelanina (26 %) en la epidermis es constante independientemente del grado de pigmentación.[19] Algunos seres humanos tienen muy poca o ninguna síntesis de melanina en sus cuerpos, una condición conocida como albinismo.[20]
Dado que la melanina es un agregado de moléculas componentes más pequeñas, existen muchos tipos diferentes de melanina con distintas proporciones y patrones de unión de estas moléculas componentes. Tanto la feomelanina como la eumelanina se encuentran en la piel y el cabello humanos, pero la eumelanina es la melanina más abundante en los seres humanos, así como la forma más propensa a ser deficiente en el albinismo.[21]
Otros organismos
editarLas melaninas tienen papeles y funciones muy diversos en varios organismos. Una forma de melanina constituye la tinta utilizada por muchos cefalópodos (véase tinta de cefalópodo) como mecanismo de defensa contra los depredadores. Las melaninas también protegen a los microorganismos, como bacterias y hongos, contra el estrés que implica el daño celular, como la radiación UV del sol y las especies reactivas del oxígeno. La melanina también protege contra los daños provocados por las altas temperaturas, las tensiones químicas (como los metales pesados y los agentes oxidantes) y las amenazas bioquímicas (como las defensas del huésped contra los microbios invasores).[22] Por lo tanto, en muchos microbios patógenos (por ejemplo, en Cryptococcus neoformans, un hongo) las melaninas parecen desempeñar papeles importantes en la virulencia y la patogenicidad al proteger al microbio contra las respuestas inmunitarias de su huésped. En los invertebrados, un aspecto importante del sistema inmunitario innato de defensa contra los patógenos invasores implica la melanina. Pocos minutos después de la infección, el microbio queda encapsulado dentro de melanina (melanización), y se cree que la generación de subproductos radicales libres durante la formación de esta cápsula ayuda a matarlos.[23] Algunos tipos de hongos, llamados hongos radiotróficos, parecen ser capaces de utilizar la melanina como pigmento fotosintético que les permite captar rayos gamma[24] y aprovechar esta energía para su crecimiento.[25]
En los peces, la melanina no sólo aparece en la piel, sino también en órganos internos como los ojos. La mayoría de las especies de peces utilizan eumelanina,[26][27] pero Stegastes apicalis y Cyprinus carpio utilizan feomelanina en su lugar.[28][29]
Las plumas más oscuras de las aves deben su color a la melanina y son menos degradadas por las bacterias que las no pigmentadas o las que contienen pigmentos carotenoides.[30] Las plumas que contienen melanina son también un 39 % más resistentes a la abrasión que las que no la contienen porque los gránulos de melanina ayudan a rellenar el espacio entre las hebras de queratina que forman las plumas.[31][32] La síntesis de la feomelanina en las aves implica el consumo de cisteína, un aminoácido semiesencial que es necesario para la síntesis del antioxidante glutatión (GSH) pero que puede ser tóxico si se encuentra en exceso en la dieta. De hecho, muchas aves carnívoras, que tienen un alto contenido proteico en su dieta, presentan una coloración basada en la feomelanina.[33]
La melanina también es importante en la pigmentación de los mamíferos.[34] El patrón del pelaje de los mamíferos está determinado por el gen agouti, que regula la distribución de la melanina.[35][36] Los mecanismos del gen se han estudiado ampliamente en ratones para comprender la diversidad de los patrones del pelaje de los mamíferos.[37]
Se ha observado que la melanina de los artrópodos se deposita en capas, produciendo así un reflector de Bragg de índice de refracción alterno. Cuando la escala de este patrón coincide con la longitud de onda de la luz visible, se produce una coloración estructural que confiere a varias especies un color iridiscente.[38][39]
Los arácnidos son uno de los pocos grupos en los que la melanina no se ha detectado fácilmente, aunque los investigadores han encontrado datos que sugieren que las arañas producen melanina.[40]
Algunas especies de polillas, incluida la polilla tigre de la madera, convierten los recursos en melanina para mejorar su termorregulación. Como la polilla tigre de la madera tiene poblaciones en un amplio rango de latitudes, se ha observado que las poblaciones más septentrionales mostraban mayores tasas de melanización. En los fenotipos macho amarillo y blanco de la polilla tigre de la madera, los individuos con más melanina tenían una mayor capacidad para atrapar el calor, pero una mayor tasa de depredación debido a una señal aposemática más débil y menos eficaz.[41]
La melanina protege a las moscas Drosophila y a los ratones contra los daños en el ADN provocados por la radiación no UV.[42] Entre los estudios importantes realizados en modelos de Drosophila se incluye el de Hopwood et al. (1985).[42] Gran parte de los conocimientos sobre los efectos radioprotectores de la melanina contra la radiación gamma proceden de los laboratorios y grupos de investigación de Irma Mosse.[43][44][45][46][47][48][49]: 1151 Mosse se inició en la radiobiología en la era soviética, recibió cada vez más financiación gubernamental a raíz del descubrimiento de microbios radiotróficos en Chernóbil y, a partir de 2022, continúa en el Instituto Bielorruso de Genética y Citología.[48] Su contribución más significativa es Mosse et al., (2000) sobre ratones,[43][44][45][46][47][48][49]: 1151 pero también incluye Mosse et al., (1994),[47] Mosse et al., (1997),[47] Mosse et al., (1998),[46] Mosse et al., (2001),[47] Mosse et al., (2002),[46][47] Mosse et al., (2006),[46][47] Mosse et al., (2007)[47] y Mosse et al., (2008).[47]
Plantas
editar
Las melaninas producidas por las plantas se denominan a veces «melaninas de catecol», ya que pueden producir catecol con la fusión alcalina. Se suele observar en el pardeamiento enzimático de frutas como el plátano. La melanina de la cáscara del castaño puede utilizarse como antioxidante y colorante.[50] La biosíntesis implica la oxidación de indol-5,6-quinona por la polifenol oxidasa de tipo tirosinasa a partir de tirosina y catecolaminas, lo que conduce a la formación de melanina catecol. A pesar de ello, muchas plantas contienen compuestos que inhiben la producción de melaninas.[51]
Interpretación como monómero único
editarActualmente se sabe que las melaninas no tienen una estructura o estequiometría única. No obstante, bases de datos químicas como PubChem incluyen fórmulas estructurales y empíricas; normalmente 3,8-Dimetil-2,7-dihidrobenzo[1,2,3-cd:4,5,6-c′d′]diindol-4,5,9,10-tetrone. Esto puede considerarse como un monómero único que explica la composición elemental medida y algunas propiedades de la melanina, pero es poco probable que se encuentre en la naturaleza.[52] Solano[52] afirma que esta tendencia engañosa proviene de un informe de una fórmula empírica en 1948,[53] pero no proporciona ningún otro detalle histórico.
Vías biosintéticas
editar| 3,8-Dimethyl-2,7-dihydrobenzo[1,2,3-cd:4,5,6-c′d′]diindole-4,5,9,10-tetrone | ||
|---|---|---|
 3,8-Dimethyl-2,7-dihydrobenzo[1,2,3-cd:4,5,6-c′d′]diindole-4,5,9,10-tetrone | ||
 3,8-Dimethyl-2,7-dihydrobenzo[1,2,3-cd:4,5,6-c′d′]diindole-4,5,9,10-tetrone, modelo de esferas y barras | ||
| Nombre IUPAC | ||
| 3,8-Dimethyl-2,7-dihydrobenzo[1,2,3-cd:4,5,6-c′d′]diindole-4,5,9,10-tetrone | ||
| General | ||
| Fórmula molecular | C18H10N2O4 | |
| Identificadores | ||
| Número CAS | 8049-97-6[54] | |
| ChEBI | 25179 | |
| ChemSpider | 4884931 | |
| PubChem | 6325610 | |
| KEGG | C01693 | |
| Propiedades físicas | ||
| Densidad | 1600 kg/m³; 1,6 g/cm³ | |
| Masa molar | 318 288 g/mol | |
| Punto de fusión | −20 °C (253 K) | |
| Punto de ebullición | 450 °C (723 K) a 550 °C (823 K) | |
| Valores en el SI y en condiciones estándar (25 ℃ y 1 atm), salvo que se indique lo contrario. | ||
El primer paso de la vía biosintética tanto de las eumelaninas como de las feomelaninas lo cataliza la tirosinasa.[55]
- Tirosina → DOPA → dopaquinona
La dopaquinona puede combinarse con la cisteína por dos vías para dar lugar a las benzotiazinas y las feomelaninas
- dopaquinona + cisteína → 5-S-cisteinildopa → intermediario benzotiazina → feomelanina.
- dopaquinona + cisteína → 2-S-cisteinildopa → intermedio benzotiazina → feomelanina.
Además, la dopaquinona puede convertirse en leucodopacromo y seguir dos vías más hasta las eumelaninas
- dopaquinona → leucodopacromo → dopacromo → ácido 5,6-dihidroxiindol-2-carboxílico → quinona → eumelanina
- dopaquinona → leucodopacromo → dopacromo → 5,6-dihidroxiindol → quinona → eumelanina.
Se pueden encontrar rutas metabólicas detalladas en la base de datos KEGG.
-
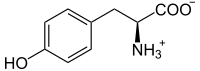 L-tirosina
L-tirosina -
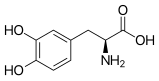 L-DOPA
L-DOPA -
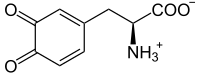 L-dopaquinona
L-dopaquinona -
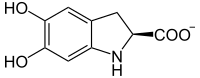 L-leucodopacromo
L-leucodopacromo -
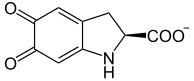 L-dopacromo
L-dopacromo

.svg)



Aspecto microscópico
editarLa melanina es marrón, no refráctil y finamente granular, con gránulos individuales de un diámetro inferior a 800 nanómetros. Esto diferencia a la melanina de los pigmentos comunes de descomposición de la sangre, que son más grandes, gruesos y refractiles, y varían en color de verde a amarillo o rojo-marrón. En las lesiones muy pigmentadas, los agregados densos de melanina pueden oscurecer los detalles histológicos. Una solución diluida de permanganato potásico es un blanqueador eficaz de la melanina.[56]
Trastornos genéticos y estados de enfermedad
editarExisten aproximadamente nueve tipos de albinismo oculocutáneo, que en la mayoría de los casos es un trastorno autosómico recesivo. Ciertas etnias tienen mayor incidencia de las distintas formas. Por ejemplo, el tipo más común, denominado albinismo oculocutáneo tipo 2 (OCA2), es especialmente frecuente entre personas de ascendencia africana negra y europeos blancos. Las personas con OCA2 suelen tener la piel clara, pero a menudo no son tan pálidas como las de OCA1. La población presenta OCA2 tienen el pelo entre rubio pálido y dorado, rubio fresa o incluso castaño y lo más habitual es que tengan los ojos azules. El 98,7-100 % de los europeos modernos son portadores del alelo derivado SLC24A5, una causa conocida de albinismo oculocutáneo no sindrómico. Se trata de un trastorno autosómico recesivo caracterizado por una reducción congénita o ausencia de pigmento de melanina en la piel, el pelo y los ojos. La frecuencia estimada de OCA2 entre los afroamericanos es de 1 entre 10 000, lo que contrasta con una frecuencia de 1 entre 36 000 en los estadounidenses blancos.[57] En algunas naciones africanas, la frecuencia del trastorno es incluso mayor, oscilando entre 1 entre 2000 y 1 entre 5000.[58] Otra forma de albinismo, el «albinismo oculocutáneo amarillo», parece ser más frecuente entre los amish, de ascendencia principalmente suiza y alemana. Las personas con esta variante IB del trastorno suelen tener el pelo y la piel blancos al nacer, pero desarrollan rápidamente una pigmentación cutánea normal en la infancia.[58]
El albinismo ocular no sólo afecta a la pigmentación de los ojos, sino también a la agudeza visual. Las personas con albinismo suelen tener una agudeza visual deficiente, entre 20/60 y 20/400. Además, dos formas de albinismo, con aproximadamente 1 de cada 2700 más prevalentes entre las personas de origen puertorriqueño, se asocian a una mortalidad superior a las muertes relacionadas con el melanoma.
La relación entre albinismo y sordera es bien conocida, aunque poco comprendida. En su tratado de 1859 El origen de las especies (On the Origin of Species), Charles Darwin observó que «los gatos que son completamente blancos y tienen los ojos azules suelen ser sordos».[59] En los humanos, la hipopigmentación y la sordera se dan juntas en el raro síndrome de Waardenburg, observado predominantemente entre los hopi en Norteamérica.[60] La incidencia del albinismo en los indios hopi se ha estimado en aproximadamente 1 de cada 200 individuos. Se han encontrado patrones similares de albinismo y sordera en otros mamíferos, incluidos perros y roedores. Sin embargo, la falta de melanina per se no parece ser directamente responsable de la sordera asociada a la hipopigmentación, ya que la mayoría de los individuos que carecen de las enzimas necesarias para sintetizar melanina tienen una función auditiva normal.[61] En cambio, la ausencia de melanocitos en la estría vascular del oído interno provoca una deficiencia coclear,[62] aunque las razones de ello no se comprenden del todo.
En la enfermedad de Parkinson, un trastorno que afecta al funcionamiento neuromotor, hay una disminución de neuromelanina en la sustancia negra y el locus coeruleus como consecuencia de la caída específica de neuronas dopaminérgicas y noradrenérgicas pigmentadas. Esto provoca una disminución de la síntesis de dopamina y norepinefrina. Aunque no se ha informado de ninguna correlación entre la raza y el nivel de neuromelanina en la sustancia negra, la incidencia significativamente menor de Parkinson en los negros que en los blancos «ha llevado a algunos a sugerir que la melanina cutánea podría servir de alguna manera para proteger la neuromelanina en la sustancia negra de las toxinas externas».[63]
Además de la deficiencia de melanina, el peso molecular del polímero de melanina puede disminuir por diversos factores como el estrés oxidativo, la exposición a la luz, la perturbación en su asociación con las proteínas de la matriz melanosomal, los cambios en el pH o en las concentraciones locales de iones metálicos. Se ha propuesto que la disminución del peso molecular o del grado de polimerización de la melanina ocular convierte el polímero, normalmente antioxidante, en prooxidante. En su estado prooxidante, se ha sugerido que la melanina está implicada en la causa y la progresión de la degeneración macular y el melanoma.[64] La rasagilina, un importante fármaco de monoterapia en la enfermedad de Parkinson, tiene propiedades de unión a la melanina y propiedades reductoras de tumores de melanoma.[65]
Sin embargo, unos niveles más altos de eumelanina también pueden ser una desventaja, además de una mayor predisposición a la deficiencia de vitamina D. La piel oscura es un factor que complica la eliminación de las nevo flámeo con láser. Los láseres son eficaces en el tratamiento de la piel blanca en general, pero tienen menos éxito en la eliminación de las manchas de vino de Oporto en personas de ascendencia asiática o africana. Las mayores concentraciones de melanina en las personas de piel más oscura simplemente difunden y absorben la radiación láser, inhibiendo la absorción de la luz por el tejido objetivo. De forma similar, la melanina puede complicar el tratamiento con láser de otras afecciones dermatológicas en personas de piel más oscura.
Las pecas y los lunares se forman donde hay una concentración localizada de melanina en la piel. Están muy asociados a la piel pálida.
La nicotina tiene afinidad por los tejidos que contienen melanina debido a su función precursora en la síntesis de melanina o a su unión irreversible a la melanina. Se ha sugerido que esto subyace a la mayor dependencia de la nicotina y a las menores tasas de abandono del tabaquismo en individuos con pigmentación más oscura.[66]
Adaptaciones humanas
editarFisiología
editarLos melanocitos insertan gránulos de melanina en vesículas celulares especializadas denominadas melanosomas. A continuación, éstos se transfieren a las células queratinocitarias de la epidermis humana. Los melanosomas de cada célula receptora se acumulan sobre el núcleo celular, donde protegen el ADN nuclear de las mutaciones causadas por la radiación ionizante de los rayos ultravioleta del sol. En general, las personas cuyos antepasados vivieron durante largos periodos en las regiones del globo cercanas al ecuador tienen mayores cantidades de eumelanina en la piel. Esto hace que sus pieles sean marrones o negras y las protege frente a altos niveles de exposición al sol, que dan lugar con más frecuencia a melanomas en las personas de piel más clara.[67]
No todos los efectos de la pigmentación son ventajosos. La pigmentación aumenta la carga térmica en climas cálidos, y las personas de piel oscura absorben un 30% más de calor de la luz solar que las de piel muy clara, aunque este factor puede verse compensado por una sudoración más profusa. En climas fríos, la piel oscura conlleva una mayor pérdida de calor por radiación. La pigmentación también dificulta la síntesis de vitamina D. Dado que la pigmentación no parece ser del todo ventajosa para la vida en los trópicos, se han propuesto otras hipótesis sobre su significado biológico; por ejemplo, un fenómeno secundario inducido por la adaptación a parásitos y enfermedades tropicales.[68]
Orígenes evolutivos
editarLos primeros humanos evolucionaron hacia un color de piel oscuro, como adaptación a la pérdida de vello corporal que aumentaba los efectos de la radiación UV. Antes de desarrollar la falta de pelo, los primeros humanos podrían haber tenido piel clara bajo el pelaje, similar a la de otros primates.[69] Los humanos anatómicamente modernos evolucionaron en África hace entre 200 000 y 100 000 años,[70] y luego poblaron el resto del mundo mediante migraciones hace entre 80 000 y 50 000 años, cruzándose en algunas zonas con ciertas especies humanas arcaicas (neandertales, denisovanos y posiblemente otras).[71] Los primeros humanos modernos tenían la piel más oscura que los actuales indígenas de África. Tras la migración y el asentamiento en Asia y Europa, la presión selectiva que ejercía la piel oscura protectora de la radiación UV disminuyó allí donde la radiación del sol era menos intensa. Esto dio lugar a la actual gama de colores de la piel humana. De las dos variantes genéticas comunes que se sabe que están asociadas a la piel humana pálida, Mc1r no parece haber sufrido selección positiva,[72] mientras que SLC24A5 sí ha sufrido selección positiva.[73]
Efectos
editarAl igual que los pueblos que han emigrado hacia el norte, los de piel clara que migran hacia el ecuador se aclimatan a la radiación solar, mucho más intensa. La naturaleza selecciona menos melanina cuando la radiación ultravioleta es débil. La piel de la mayoría de las personas se oscurece cuando se exponen a la luz ultravioleta, lo que les proporciona más protección cuando la necesitan. Éste es el propósito fisiológico del bronceado. Las personas de piel oscura, que producen más eumelanina protectora de la piel, tienen una mayor protección contra las quemaduras solares y el desarrollo de melanoma, una forma potencialmente mortal de cáncer de piel, así como otros problemas de salud relacionados con la exposición a radiaciones solares intensas, incluida la fotodegradación de ciertas vitaminas como las riboflavinas, los carotenoides, el tocoferol y el folato.[74]
La melanina de los ojos, en el iris y la coroides, ayuda a proteger de la luz ultravioleta y la luz visible de alta frecuencia; las personas con ojos azules, verdes y grises tienen más riesgo de sufrir problemas oculares relacionados con el sol. Además, el cristalino amarillea con la edad, proporcionando una protección añadida. Sin embargo, el cristalino también se vuelve más rígido con la edad, perdiendo la mayor parte de su capacidad de acomodación —la capacidad de cambiar de forma para enfocar de lejos a cerca—, un perjuicio debido probablemente al entrecruzamiento de proteínas causado por la exposición a los rayos UV.
Investigaciones recientes sugieren que la melanina puede desempeñar una función protectora distinta de la fotoprotección.[75] La melanina es capaz de quelar eficazmente iones metálicos a través de sus grupos carboxilato e hidroxilo fenólico, a menudo con mucha más eficacia que el potente ligando quelante etilendiaminotetraacetato (EDTA). Así, puede servir para secuestrar iones metálicos potencialmente tóxicos, protegiendo al resto de la célula. Esta hipótesis se ve respaldada por el hecho de que la pérdida de neuromelanina, observada en la enfermedad de Parkinson, va acompañada de un aumento de los niveles de hierro en el cerebro.
Propiedades físicas y aplicaciones tecnológicas
editarExisten pruebas de la existencia de un heteropolímero altamente reticulado unido covalentemente a las melanoproteínas de andamiaje de la matriz.[76] Se ha propuesto que la capacidad de la melanina para actuar como antioxidante es directamente proporcional a su grado de polimerización o peso molecular.[77] Unas condiciones subóptimas para la polimerización efectiva de los monómeros de melanina pueden conducir a la formación de melanina prooxidante de menor peso molecular, implicada en la causalidad y progresión de la degeneración macular y el melanoma.[78] Las vías de señalización que aumentan la regulación de la melanización en el epitelio pigmentario de la retina (EPR) también pueden estar implicadas en la desregulación de la fagocitosis del segmento externo del bastón por el EPR. Este fenómeno se ha atribuido en parte a la preservación foveal en la degeneración macular.[79]
Papel en la metástasis del melanoma
editarLas células de melanoma muy pigmentadas tienen un módulo de Young de aproximadamente 4,93 kPa, en comparación con las células no pigmentadas, con un valor de 0,98 kPa.[80] La elasticidad de las células de melanoma es crucial para la metástasis y el crecimiento; los tumores no pigmentados eran más grandes que los pigmentados y se propagaban con mucha más facilidad. Tanto las células pigmentadas como las no pigmentadas están presentes en los tumores de melanoma, por lo que ambos pueden ser resistentes a los fármacos y metastásicos.[80]
Véase también
editarReferencias
editar- ↑ Casadevall, Arturo (2018). «Melanin triggers antifungal defences». Nature (en inglés) 555 (7696): 319-320. Bibcode:2018Natur.555..319C. ISSN 0028-0836. PMID 29542711. doi:10.1038/d41586-018-02370-x.
- ↑ a b Cao, Wei; Zhou, Xuhao; McCallum, Naneki C.; Hu, Ziying; Ni, Qing Zhe; Kapoor, Utkarsh; Heil, Christian M.; Cay, Kristine S. et al. (9 de febrero de 2021). «Unraveling the Structure and Function of Melanin through Synthesis». Journal of the American Chemical Society (en inglés) 143 (7): 2622-2637. ISSN 0002-7863. PMID 33560127. doi:10.1021/jacs.0c12322.
- ↑ Haining, Robert L.; Achat-Mendes, Cindy (Marzo de 2017). «Neuromelanin, one of the most overlooked molecules in modern medicine, is not a spectator». Neural Regeneration Research (en inglés) 12 (372–375). PMC 5399705. PMID 28469642. doi:10.4103/1673-5374.202928.
- ↑ Meredith, P.; Riesz, J. (2004). «Radiative relaxation quantum yields for synthetic eumelanin». Photochemistry and Photobiology (en inglés) 79 (2): 211-6. PMID 15068035. doi:10.1111/j.1751-1097.2004.tb00012.x.
- ↑ Brenner, M.; Hearing, V. J. (2008). «The protective role of melanin against UV damage in human skin». Photochemistry and Photobiology (en inglés) 84 (3): 539-49. PMC 2671032. PMID 18435612. doi:10.1111/j.1751-1097.2007.00226.x.
- ↑ a b c d Alaluf, Simon; Heath, Alan; Carter, Nik; Atkins, Derek; Mahalingam, Harish; Barrett, Karen; Kolb, Ria; Smit, Nico (2001). «Variation in Melanin Content and Composition in Type V and VI Photoexposed and Photoprotected Human Skin: The Dominant Role of DHI». Pigment Cell Research (en inglés) 14 (5): 337-347. ISSN 0893-5785. PMID 11601655. doi:10.1034/j.1600-0749.2001.140505.x.
- ↑ Ito, S.; Wakamatsu, K. (diciembre de 2011). «Diversity of human hair pigmentation as studied by chemical analysis of eumelanin and pheomelanin». Journal of the European Academy of Dermatology and Venereology (en inglés) 25 (12): 1369-1380. ISSN 1468-3083. PMID 22077870. doi:10.1111/j.1468-3083.2011.04278.x.
- ↑ Slominski, A.; Tobin, D. J.; Shibahara,, S.; Wortsman, J. (octubre de 2004). «Melanin pigmentation in mammalian skin and its hormonal regulation». Physiological Reviews (en inglés) 84 (4): 1155-228. PMID 15383650. doi:10.1152/physrev.00044.2003.
- ↑ «pheomelanin». MetaCyc Metabolic Pathway Database (en inglés). 2010.
- ↑ Thody, A. J.; Higgins, E. M.; Wakamatsu, K.; Ito, S.; Burchill, S. A.; Marks, J. M. (agosto de 1991). «Pheomelanin as well as eumelanin is present in human epidermis». The Journal of Investigative Dermatology (en inglés) 97 (2): 340-344. PMID 2071942. doi:10.1111/1523-1747.ep12480680.
- ↑ Greco G; Panzella, L.; Napolitano, A.; Verotta, L.; d'Ischia, M. (abril de 2011). «Uncovering the structure of human red hair pheomelanin: benzothiazolylthiazinodihydroisoquinolines as key building blocks». Journal of Natural Products (en inglés) 74 (4): 675-82. PMID 21341762. doi:10.1021/np100740n.
- ↑ Fedorow, H.; Tribl, F.; Halliday, G.; Gerlach, M.; Riederer, P.; Double, K. L. (2005). «Neuromelanin in human dopamine neurons: comparison with peripheral melanins and relevance to Parkinson's disease». Prog Neurobiol (en inglés) 75 (2): 109-124. PMID 15784302. doi:10.1016/j.pneurobio.2005.02.001.
- ↑ Double, K. L. (2006). «Functional effects of neuromelanin and synthetic melanin in model systems». J Neural Transm (en inglés) 113 (6): 751-756. PMID 16755379. doi:10.1007/s00702-006-0450-5.
- ↑ Kolipakala, Rakshita Sukruth; Basu, Suranjana; Sarkar, Senjuti; Biju, Beneta Merin; Salazar, Daniela; Reddy, Likhit et al. (7 de agosto de 2024). «Fungal Peptidomelanin: A Novel Biopolymer for the Chelation of Heavy Metals». ACS Omega (en inglés). ISSN 2470-1343. PMC 11359623. doi:10.1021/acsomega.4c03704.
- ↑ Wei, Cao; et al (2020). «Selenomelanin: An Abiotic Selenium Analogue of Pheomelanin». Journal of the American Chemical Society (en inglés) 142 (29): 12802-12810. PMID 32638590. doi:10.1021/jacs.0c05573.
- ↑ Mark Heiden (8 de julio de 2020). «New biomaterial could shield against harmful radiation» (en inglés). Universidad Northwestern.
- ↑ Prota, G.; Searle, A. G. (1978). «Biochemical sites of gene action for melanogenesis in mammals». Annales de Génétique et de Sélection Animale (en inglés) 10 (1): 1-8. PMC 2757330. PMID 22896083. doi:10.1186/1297-9686-10-1-1.
- ↑ Solano, F. (2014). «Melanins: Skin Pigments and Much More—Types, Structural Models, Biological Functions, and Formation Routes». New Journal of Science (en inglés): 1-28. doi:10.1155/2014/498276.
- ↑ Del Bino, Sandra; Ito, Shosuke; Sok, Juliette; Wakamatsu, Kazumasa (2022). «5,6‐Dihydroxyindole eumelanin content in human skin with varying degrees of constitutive pigmentation». Pigment Cell & Melanoma Research (en inglés) 35 (6): 622-626. ISSN 1755-1471. PMC 9804219. PMID 35933709. doi:10.1111/pcmr.13062.
- ↑ Cichorek, Mirosława; Wachulska, Małgorzata; Stasiewicz, Aneta; Tymińska, Agata (20 de febrero de 2013). «Skin melanocytes: biology and development». Advances in Dermatology and Allergology (en inglés) 30 (1): 30-41. PMC 3834696. PMID 24278043. doi:10.5114/pdia.2013.33376.
- ↑ «oculocutaneous albinism». Genetics Home Reference (en inglés). Consultado el 25 de septiembre de 2017.
- ↑ Hamilton, A. J.; Gomez, B. L. (marzo de 2002). «Melanins in fungal pathogens». Journal of Medical Microbiology (en inglés) 51 (3): 189-91. PMID 11871612. doi:10.1099/0022-1317-51-3-189.
- ↑ Cerenius, L.; Söderhäll, K. «The prophenoloxidase-activating system in invertebrates». Immunological Reviews (en inglés) 198 (116–26). PMID 15199959. doi:10.1111/j.0105-2896.2004.00116.x.
- ↑ Castelvecchi, Davide (26 de mayo de 2007). «Dark Power: Pigment seems to put radiation to good use». Science News (en inglés) 171 (21): 325. doi:10.1002/scin.2007.5591712106.
- ↑ Dadachova, E.; Bryan, R. A.; Huang, X.; et al (2007). «Ionizing radiation changes the electronic properties of melanin and enhances the growth of melanized fungi». PLOS ONE (en inglés) 2 (5): e457. Bibcode:2007PLoSO...2..457D. PMC 1866175. PMID 17520016. doi:10.1371/journal.pone.0000457.
- ↑ Sköld, H. N.; Aspengren, S.; Cheney, K. L.; Wallin, M. (2016). «Chapter Four - Fish Chromatophores—From Molecular Motors to Animal Behavior». International Review of Cell and Molecular Biology (en inglés) 321: 171-219. PMID 26811288. doi:10.1016/bs.ircmb.2015.09.005.
- ↑ Kottler, Verena A.; Künstner, Axel; Schartl, Manfred (may de 2015). «Pheomelanin in fish?». Pigment Cell & Melanoma Research (en inglés) 28 (3): 355-356. ISSN 1755-1471. PMID 25660115. doi:10.1111/pcmr.12359.
- ↑ Xu, Peng; Zhang, Xiaofeng; Wang, Xumin; Li, Jiongtang; Liu, Guiming; Kuang, Youyi; Xu, Jian; Zheng, Xianhu et al. (noviembre de 2014). «Genome sequence and genetic diversity of the common carp, Cyprinus carpio». Nature Genetics (en inglés) 46 (11): 1212-1219. ISSN 1061-4036. PMID 25240282. doi:10.1038/ng.3098.
- ↑ Mouchet SR, Cortesi F, Bokic B, Lazovic V, Vukusic P, Marshall NJ, Kolaric B (1 de noviembre de 2023). «Morphological and Optical Modification of Melanosomes in Fish Integuments upon Oxidation». Optics (en inglés) 4 (4): 563-562. doi:10.3390/opt4040041.
- ↑ Gunderson, Alex R.; Frame, Alicia M. (1 de septiembre de 2008). «Resistance of melanized feathers to bacterial degradation: is it really so black and white?». Journal of Avian Biology (en inglés) 39 (5): 539-545. doi:10.1111/j.0908-8857.2008.04413.x.
- ↑ Bonser, Richard H. C. (1995). «Melanin and the Abrasion Resistance of Feathers». Condor (en inglés) 97 (2): 590-591. doi:10.2307/1369048.
- ↑ Galván, Ismael; Solano, Francisco (8 de abril de 2016). «Bird Integumentary Melanins: Biosynthesis, Forms, Function and Evolution». International Journal of Molecular Sciences (en inglés) 17 (4): 520. PMC 4848976. PMID 27070583. doi:10.3390/ijms17040520.
- ↑ Rodríguez-Martínez, Sol; Galván, Ismael (2020). «Juvenile pheomelanin-based plumage coloration has evolved more frequently in carnivorous species». Ibis (en inglés) 162 (1): 238-244. ISSN 1474-919X. doi:10.1111/ibi.12770.
- ↑ Jimbow, K.; Quevedo W. C., Jr.; Fitzpatrick, T. B.; Szabo, G. (julio de 1976). «Some aspects of melanin biology: 1950–1975». The Journal of Investigative Dermatology (en inglés) 67 (1): 72-89. PMID 819593. doi:10.1111/1523-1747.ep12512500.
- ↑ Meneely, Philip (2014). Genetic Analysis: Genes, Genomes, and Networks in Eukaryotes (en inglés). Oxford University Press. ISBN 9780199681266.
- ↑ Griffiths, Anthony JF; Miller, Jeffrey H.; Suzuki, David T.; Lewontin, Richard C.; Gelbart, William M. (2000). «Gene interaction in coat color of mammals». NCBI.
- ↑ Millar, S. E.; Miller, M. W.; Stevens, M. E.; Barsh, G. S. (octubre de 1995). «Expression and transgenic studies of the mouse agouti gene provide insight into the mechanisms by which mammalian coat color patterns are generated». Development (en inglés) 121 (10): 3223-3232. PMID 7588057. doi:10.1242/dev.121.10.3223.
- ↑ Neville, A. C. (2012). Biology of the Arthropod Cuticle (en inglés). Springer Science & Business Media. ISBN 9783642809101.
- ↑ Mouchet, Sébastien R. (2021). Natural Photonics and Bioinspiration (en inglés) (1ª edición). Artech House. ISBN 978-163-081-797-8.
- ↑ Hsiung, B.-K.; Blackledge, T. A. (2015). «Spiders do have melanin after all». Journal of Experimental Biology (en inglés) 218 (22). PMID 26449977. doi:10.1242/jeb.128801.
- ↑ Hegna, Robert H.; Nokelainen, Ossi; Hegna, Jonathan R.; Mappes, Johanna. «To quiver or to shiver: increased melanization benefits thermoregulation, but reduces warning signal efficacy in the wood tiger moth». Proc. R. Soc. B (en inglés) 280 (1755): 20122812. PMC 3574392. PMID 23363631. doi:10.1098/rspb.2012.2812.
- ↑ a b Mosse, Irma B.; Dubovic, Boris V.; Plotnikova, Svetlana I.; Kostrova, Ludmila N.; Molophei, Vadim; Subbot, Svetlana T.; Maksimenya, Inna P. (20-25 de mayo de 2001). «Melanin is Effective Radioprotector against Chronic Irradiation and Low Radiation Doses». En Obelic, B.; Ranogajev-Komor, M.; Miljanic, S. et al., eds. IRPA Regional Congress on Radiation Protection in Central Europe: Radiation Protection and HealthINIS (Dubrovnik (Croatia): Asociación Croata de Protección Radiológica): 35 (de 268).
- ↑ a b Gessler, N. N.; Egorova, A. S.; Belozerskaya, T. A. (2014). «Melanin pigments of fungi under extreme environmental conditions (Review)». Applied Biochemistry and Microbiology (en inglés) (Pleiades Publishing) 50 (2): 105-113. ISSN 0003-6838. PMID 25272728. doi:10.1134/s0003683814020094.
- ↑ a b Nenoi, M.; Wang, B.; Vares, G. (12 de junio de 2014). «In vivo radioadaptive response». Toxicology (en e) (Sage) 34 (3): 272-283. ISSN 0960-3271. PMC 4442823. PMID 24925363. doi:10.1177/0960327114537537.
- ↑ a b Liu, Heng; Yang, Youyuan; Liu, Yu; Pan, Jingjing; Wang, Junqing; Man, Fengyuan; Zhang, Weiguo; Liu, Gang (7 de febrero de 2020). «Melanin-Like Nanomaterials for Advanced Biomedical Applications: A Versatile Platform with Extraordinary Promise». Advanced Science (en inglés) (Wiley-VCH) 7 (7): 1903129. ISSN 2198-3844. PMC 7141020. PMID 32274309. doi:10.1002/advs.201903129.
- ↑ a b c d e Mosse, Irma B. (2012). «Genetic effects of ionizing radiation – some questions with no answers». Journal of Environmental Radioactivity (en inglés) (Elsevier) 112: 70-75. Bibcode:2012JEnvR.112...70M. ISSN 0265-931X. PMID 22683898. doi:10.1016/j.jenvrad.2012.05.009.
- ↑ a b c d e f g h i Mosse, Irma; Kilchevsky, Alexander; Kilchevsky, Alexander; Nikolova, Nevena; Zhelev, Nikolai (14 de diciembre de 2016). «Some problems and errors in cytogenetic biodosimetry». Biotechnology & Biotechnological Equipment (en inglés) (Taylor & Francis) 31 (3): 460-468. ISSN 1310-2818. doi:10.1080/13102818.2016.1259018.
- ↑ a b c Mosse, Irma (18 de enero de 2022). «Radiobiology in my life – Irma Mosse». International Journal of Radiation Biology (en inglés) (Taylor & Francis) 98 (3: Women in Radiobiology): 474-478. ISSN 0955-3002. PMID 34994663. doi:10.1080/09553002.2022.2026517.
- ↑ a b Dadachova, Ekaterina; Casadevall, Arturo (2011). Horikoshi, Kōki, ed. Extremophiles handbook (en inglés). Tokio/Ciudad de Nueva York: Springer. ISBN 978-4-431-53898-1. OCLC 700199222.
- ↑ Yao, Zeng-Yu; Qi, Jian-Hua (22 de abril de 2016). «Comparison of Antioxidant Activities of Melanin Fractions from Chestnut Shell». Molecules (en inglés) 21 (4): 487. PMC 6273334. PMID 27110763. doi:10.3390/molecules21040487.
- ↑ Kim, Y.-J.; Uyama, H. (15 de mayo de 2005). «Tyrosinase inhibitors from natural and synthetic sources: structure, inhibition mechanism and perspective for the future». Cellular and Molecular Life Sciences (en inglés) 62 (15): 1707-1723. PMC 11139184. PMID 15968468. doi:10.1007/s00018-005-5054-y.
- ↑ a b Solano, F. (2014). «Melanins: Skin Pigments and Much More—Types, Structural Models, Biological Functions, and Formation Routes». New Journal of Science (en inglés) 2014 (498276): 1-28. doi:10.1155/2014/498276.
- ↑ Mason, H. S. (1948). «The chemistry of melanin. Mechanism of the oxidation of dihydroxyphenylalanine by tyrosinase». Journal of Biological Chemistry (en inglés) 172 (1): 83-99. PMID 18920770. doi:10.1016/S0021-9258(18)35614-X.
- ↑ Número CAS
- ↑ Zaidi, Kamal Uddin; Ali, Ayesha S.; Ali, Sharique A.; Naaz, Ishrat (2014). «Microbial Tyrosinases: Promising Enzymes for Pharmaceutical, Food Bioprocessing, and Environmental Industry». Biochemistry Research International (en inglés) 2014: 1-16 (ver Fig. 3). PMC 4033337. PMID 24895537. doi:10.1155/2014/854687.
- ↑ «Melanin». pubchem.ncbi.nlm.nih.gov (en inglés). Consultado el 25 de septiembre de 2017.
- ↑ «Oculocutaneous Albinism» (en inglés). Archivado desde el original el 23 de diciembre de 2008.
- ↑ a b Peracha, Mohammed O.; Cosgrove, Frances M.; Garcia-Valenzuela, Enrique; Eliott, Dean (5 de noviembre de 2015). «Ocular Manifestations of Albinism: Background, Pathophysiology, Epidemiology». En Roy, Sr. Hampton; Talavera, Francisco; Rowsey, J. James, ed. Medscape (en inglés). WebMD. Archivado desde el original el 28 de marzo de 2017. Consultado el 8 de septiembre de 2022.
- ↑ «Causes of Variability» (en inglés). Archivado desde el original el 23 de septiembre de 2006. Consultado el 18 de septiembre de 2006.
- ↑ EntrezGene 300700
- ↑ EntrezGene 606933
- ↑ Cable, J.; Huszar, D.; Jaenisch, R.; Steel, K. P. (febrero de 1994). «Effects of mutations at the W locus (c-kit) on inner ear pigmentation and function in the mouse». Pigment Cell Research (en inglés) 7 (1): 17-32. PMID 7521050. doi:10.1111/j.1600-0749.1994.tb00015.x.
- ↑ Tamara A. Smith; Richard A. Prayson. «Lewy Body Disease» (en inglés). Archivado desde el original el 21 de julio de 2009.
- ↑ Meyskens, F. L.; Farmer, P.; Fruehauf, J. P. (junio de 2001). «Redox regulation in human melanocytes and melanoma». Pigment Cell Research (en inglés) 14 (3): 148-54. PMID 11434561. doi:10.1034/j.1600-0749.2001.140303.x.
- ↑ Meier-Davis, D. R.; Dines, K.; Arjmand, F. M.; et al (diciembre de2012). «Comparison of oral and transdermal administration of rasagiline mesylate on human melanoma tumor growth in vivo». Cutaneous and Ocular Toxicology (en inglés) 31 (4): 312-7. PMID 22515841. doi:10.3109/15569527.2012.676119.
- ↑ King, G.; Yerger, V. B.; Whembolua, G. L.; Bendel, R. B.; Kittles, R.; Moolchan, E. T. (junio de 2009). «Link between facultative melanin and tobacco use among African Americans». Pharmacology Biochemistry and Behavior (en inglés) 92 (4): 589-96. PMID 19268687. doi:10.1016/j.pbb.2009.02.011.
- ↑ «Human Skin Color Variation». The Smithsonian Institution's Human Origins Program (en inglés). 20 de junio de 2012. Consultado el 24 de agosto de 2019.
- ↑ Berth-Jones, J. (2010). «Constitutive pigmentation, human pigmentation and the response to sun exposure». En Tony Burns; Stephen Breathnach; Neil Cox; Christopher Griffiths, ed. Rook's Textbook of Dermatology (en inglés) 3 (8ª edición). Wiley-Blackwell. p. 58.9. ISBN 978-1-4051-6169-5.
- ↑ Wade, Nicholas. «Why Humans and Their Fur Parted Ways». The New York Times (en inglés). ISSN 0362-4331. Consultado el 19 de agosto de 2003.
- ↑ Tishkoff, S. A.; Reed, F. A.; Friedlaender, F. R., et al. (mayo 2009). «The genetic structure and history of Africans and African Americans». Science (en inglés) 324 (5930): 1035-44. PMC 2947357. PMID 19407144. doi:10.1126/science.1172257.
- ↑ Carl Zimmer (22 de septiembre de 2016). «A Single Migration From Africa Populated the World, Studies Find». The New York Times (en inglés).
- ↑ Harding, R. M.; Healy, E.; Ray, A. J. et al. (abril del 2000). «Evidence for variable selective pressures at MC1R». American Journal of Human Genetics (en inglés) 66 (4): 1351-61. PMC 1288200. PMID 10733465. doi:10.1086/302863.
- ↑ Lamason, R. L.; Mohideen, M. A.; Mest, J. R., etal (diciembre de 2005). «SLC24A5, a putative cation exchanger, affects pigmentation in zebrafish and humans». Science (en inglés) 310 (5755): 1782-6. Bibcode:2005Sci...310.1782L. PMID 16357253. doi:10.1126/science.1116238.
- ↑ Jablonski, Nina G.; Chaplin, George (11 de mayo de 2010). «Human skin pigmentation as an adaptation to UV radiation». Proceedings of the National Academy of Sciences (en inglés) 107 (Supplement 2): 8962-8968. Bibcode:2010PNAS..107.8962J. PMC 3024016. PMID 20445093. doi:10.1073/pnas.0914628107.
- ↑ Liu, Y.; Hong, L.; Kempf, V. R.; Wakamatsu, K; Ito, S.; Simon, J. D. (junio de 2004). «Ion-exchange and adsorption of Fe(III) by Sepia melanin». Pigment Cell Research (en inglés) 17 (3): 262-9. PMID 15140071. doi:10.1111/j.1600-0749.2004.00140.x.
- ↑ Donatien, PD; Orlow, SJ (agosto de 1995). «Interaction of melanosomal proteins with melanin». European Journal of Biochemistry (en inglés) 232 (1): 159-64. PMID 7556145. doi:10.1111/j.1432-1033.1995.tb20794.x.
- ↑ Sarangarajan, R.; Apte, S. P. (2005). «Melanin aggregation and polymerization: possible implications in age-related macular degeneration». Ophthalmic Research (en inglés) 37 (3): 136-41. PMID 15867475. doi:10.1159/000085533.
- ↑ Meyskens, F. L.; Farmer, P. J.; Anton-Culver, H. (abril de 2004). «Etiologic pathogenesis of melanoma: a unifying hypothesis for the missing attributable risk». Clinical Cancer Research (en inglés) 10 (8): 2581-3. PMID 15102657. doi:10.1158/1078-0432.ccr-03-0638.
- ↑ Sarangarajan, R.; Apte, S. P. (2005). «Melanization and phagocytosis: implications for age related macular degeneration». Molecular Vision (en inglés) 11: 482-90. PMID 16030499.
- ↑ a b Sarna, Michal; Krzykawska-Serda, Martyna; Jakubowska, Monika; Zadlo, Andrzej; Urbanska, Krystyna (26 de junio de 2019). «Melanin presence inhibits melanoma cell spread in mice in a unique mechanical fashion». Scientific Reports (en inglés) 9 (1). Bibcode:2019NatSR...9.9280S. ISSN 2045-2322. PMC 6594928. PMID 31243305. doi:10.1038/s41598-019-45643-9.
Enlaces externos
editar Wikimedia Commons alberga una categoría multimedia sobre Melanina.
Wikimedia Commons alberga una categoría multimedia sobre Melanina. Wikcionario tiene definiciones y otra información sobre melanina.
Wikcionario tiene definiciones y otra información sobre melanina.
- «Absorption spectrum of melanin.» (en inglés). Departamento de Informática y Tecnología.
- «Tyrosine metabolism—Reference pathway». Kyoto Encyclopedia of Genes and Genomes (en inglés). Consultado el 13 de junio de 2024.
- «Melanogenesis». Kyoto Encyclopedia of Genes and Genomes (en inglés). Consultado el 13 de junio de 2024.