Cnidaria
Los cnidarios (Cnidaria, del griego ''kníde'', ortiga) (pronunciación: [nidarios]) son un filo de animales diblásticos relativamente simples, que viven exclusivamente en ambientes acuáticos, mayoritariamente marinos. Agrupa alrededor de 10 000 especies, englobando medusas, pólipos, corales, anémonas e hidras (únicos cnidarios que habitan en agua dulce).
| Cnidaria | ||
|---|---|---|
| Rango temporal: Ediacárico – Reciente | ||
 | ||
| Taxonomía | ||
| Dominio: | Eukaryota | |
| Reino: | Animalia | |
| Subreino: | Eumetazoa | |
| (sin rango) | ParaHoxozoa | |
| Filo: |
Cnidaria Verrill 1865, Hatschek 1888 | |
| Subfilos y clases | ||
| ||


El nombre del filo alude a una característica diagnóstica propia de estos animales, la presencia de unas células urticantes llamadas cnidoblastos,[1] presentes en los tentáculos y boca de todos los miembros del filo; estos funcionan como cavidad y se encargan de guardar los cnidocitos, que son cápsulas en donde es almacenado el veneno y son lanzadas mediante un filamento hacia la presa, enganchándola con un aguijón para poder liberar las toxinas. Tienen simetría radial y su plan corporal es en forma de saco. Son los animales más simples que presentan células nerviosas y órganos de los sentidos (estatocistos, ocelos).
Son un grupo antiguo, con una larga historia fósil que se remonta, probablemente, a la fauna del Precámbrico, hace unos 700 millones de años. No obstante, análisis genéticos del reloj molecular de sus mitocondrias sugieren una edad muy anterior para el grupo corona de todos los cnidarios, estimada en unos 741 millones de años, mucho antes de que haya fósiles que hayan perdurado hasta la actualidad.[2]
Características generales
editarLos cnidarios son metazoarios diblásticos, es decir, son animales sin tejidos que poseen dos capas embrionarias (ectodermo y endodermo); aunque a veces existe un tejido análogo (ectomesodermo) de origen ectodérmico, no de origen endodérmico como el auténtico mesodermo de los triblásticos, y del cual nunca derivan órganos internos complejos. Sus células se organizan en dos capas que actúan como unidades funcionales (tejidos), aunque muchas células todavía guardan cierta independencia y cierta totipotencia.
Estos organismos pueden ser individuos solitarios o coloniales y sésiles, sedentarios o libres nadadores. Algunos de ellos son suspensívoros, sobre todo las especies sésiles, pero la mayoría son carnívoros,[3] por lo que tienen que cazar. Para llevar a cabo la caza de alimento, estos invertebrados poseen células especializadas llamadas cnidoblastos,[1] ubicados en sus tentáculos y boca; funcionan como cavidad en donde se encuentran guardados los cnidocitos, que son cápsulas en donde se almacena el veneno y son lanzadas mediante un filamento hacia la presa, enganchándola con un aguijón para poder liberar las toxinas. Estas células son muy sensibles, por lo que basta con rozar los tentáculos del animal para que sus aguijones sean expulsados. En el esquema de abajo se puede observar un cnidoblasto y el proceso de la descarga de un cnidocito.

Su organización corporal es en forma de saco; el aparato digestivo tiene un solo orificio que actúa como boca y ano al mismo tiempo, y se conoce como cavidad gastrovascular, en donde se realiza la digestión, se utiliza también como sistema de distribución de nutrientes y del oxígeno, y como sistema excretor.
La pared del cuerpo consta de:
- Ectodermis. Homóloga del ectodermo. Es la capa externa. Función protectora.
- Gastrodermis. Homóloga del endodermo. Es la capa interna; delimita una amplia cavidad, la única del animal, llamada cavidad gastrovascular, que comunica por un único orificio con el exterior que se usa para la alimentación, la excreción e incluso la expulsión de los productos genitales.
- Mesoglea. Entre ambas capas se encuentra la mesoglea, que puede ser muy fina y acelular, o estar bien desarrollada y tener distintos tipos celulares (según el grupo).
Tienen uno o varios tentáculos alrededor de la boca. El sistema nervioso forma de una red o plexo; en muchos grupos hay protoneuronas no polarizadas, aunque también puede haber neuronas polarizadas, células sensoriales e incluso agrupación de las mismas en órganos sensoriales.
Tienden al polimorfismo, en especial formas coloniales. No hay aparato excretor, aparato respiratorio, ni aparato circulatorio. Estas funciones se realizan a través de la cavidad gastrovascular o de la ectodermis.
Son esencialmente marinos (99%). El resto son de agua dulce, como la hidra, o ciertas medusas de grandes lagos africanos como la medusa Craspedacusta. Son siempre acuáticos.
Pueden vivir de forma individual o en colonias, fijados al sustrato o libres, incluso los hay nadadores. En ocasiones, parte de las fases son planctónicas (móviles, pero arrastrados por las corrientes).
Se conocen aproximadamente 10 000 especies, de tamaño variable, de 1-2 mm hasta 1 m de diámetro en algunas medusas, o hasta 3 m de diámetro en algunos pólipos.
El color también es variable aunque muchas formas del plancton son transparentes. Otras son coloreadas, y presentan prácticamente todos los colores.
Morfología
editar
Sin atender a las peculiaridades de cada grupo aparecen en dos formas fundamentales:
Forma sésil: Pólipo
editar
En términos generales (ver por grupos para más información), es un animal de paredes finas y amplia cavidad gastrovascular. Mesoglea muy poco desarrollada. Cuerpo más o menos columnar. Por el lado aboral (ver simetría radial) se relaciona con el sustrato. De adultos, pueden ser sésiles o libres, solitarios o coloniales.
Pueden ser dioicos o hermafroditas, y tienen una reproducción sexual y otra asexual.
Tamaño variable:
- Halamohydra: de la meiofauna, muy pequeña
- Microhydra: pequeña y sin tentáculos. Es la fase pólipo de la medusa Craspedacusta. De 1-2 mm.
- Branchiocerianthus: fondos fangosos, mucha profundidad. Hasta 3 m.
-
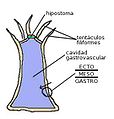 Diagrama general de un pólipo (sin atender a grupos)
Diagrama general de un pólipo (sin atender a grupos) -
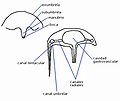 Diagrama general de una medusa (sin atender a grupos)
Diagrama general de una medusa (sin atender a grupos)


Forma libre: Medusa
editar
1.- Ectodermis; 2.- Mesoglea; 3.- Gastrodermis; 4.- Estómago; 5.- Canal radial; 6.- Canal circular; 7.- Tentáculo; 8.- Velo; 9.- Anillo nervioso externo; 10.- Anillo nervioso interno; 11.- Gónada; 12.- Manubrio; 13.- Boca; 14.- Exumbrela; 15.- Subumbrela..
Presenta dos superficies perfectamente definidas; una cóncava donde se sitúa la boca (lado oral, véase simetría radial) y otra la opuesta (aboral). Al conjunto se le conoce como umbrela, donde se diferencian la exumbrela (aboral) y subumbrela (oral).
La boca se prolonga en un manubrio. Del manubrio pueden salir (o no, según los grupos) tentáculos. Además, la umbrela puede alargarse en tentáculos umbrelares (que pueden o no presentar cavidad tentacular, una extensión de la cavidad gastrovascular).
La boca desemboca en la cavidad gastrovascular de la que parten canales radiales hacia las paredes de la medusa. El canal umbrelar da la vuelta a toda la medusa. Si los tentáculos son huecos, también aparece el canal tentacular.
Toda la cavidad (también los canales) está tapizada por gastrodermis. El resto por ectodermis. Hay mesoglea muy desarrollada en la exumbrela, mientras que en la subumbrela está muy poco desarrollada.
Tamaño variable, desde pequeñas medusas de pólipos coloniales como Obelia (1-2 mm) hasta grandes medusas como la nomura que puede llegar a 2 m de diámetro y pesar hasta 220 kilogramos. Las medusas gigantes son animales míticos.
Anatomía interna
editarVeamos los tipos celulares que aparecen en los cnidarios:
Ectodermis
editar
La ectodermis consta de diversos tipos de células:
- Células mioepiteliales o epiteliomusculares. El tipo fundamental es una célula (medusa y pólipo) cilíndrica con dos partes definidas, una epitelial apical y otra basal contráctil que posee filamentos de actina y miosina. Tienen, pues, doble función, revestimiento y contráctil.
- Células intersticiales o basales. Están en la base de las células mioepiteliales, dispuestas a modo de cuña, y tienen un núcleo muy grande. Son muy ricas en ribosomas, y son células totipotentes, de modo que pueden originar cualquier otro tipo celular.
- Cnidocitos o cnidoblastos. Los cnidocitos están esparcidos por toda la epidermis, alojados o invaginados en las células mioepiteliales; especialmente en los tentáculos. Se trata de un tipo muy especializado de célula, exclusiva de los cnidarios. Tienen un cilio rígido (cnidocilio) que es un receptor de estímulo; en la parte basal tiene prolongaciones que contactan con otras células o con el sistema nervioso (cnidopodios); en el interior de esta célula hay una gran vacuola (cnido o nematocisto), dentro de la cual se encuentra el filamento urticante (el disparo de este filamento es la "picadura" de las medusas, pólipos, anémonas, etc).
- Neuronas. Células nerviosas, unas sin polarizar (protoneuronas) y otras polarizadas, que pueden ser bipolares o multipolares. El sistema nervioso forma un plexo que se sitúa bajo la epidermis.
- Células sensoriales. Células alargadas con el extremo apical redondeado o provisto de un cilio sensible y la zona basal provista de una o varias prolongaciones que conectan con las neuronas
No está claro si las células mioepiteliales descansan sobre una verdadera lámina basal, como los tejidos epiteliales auténticos de otros metazoos, aunque puede interpretarse que la propia mesoglea es la lámina basal del epitelio de los cnidarios.[4]
Mesoglea
editarSustancia gelatinosa que separa las dos capas epiteliales. Puede variar de una membrana delgada, no celular, hasta una gruesa capa gelatinosa con amebocitos errantes o sin ellos. Presenta abundante colágeno y precolágeno. Contiene numerosas células, como escleroblastos.[5]
Gastrodermis
editar- Células nutritivomusculares o mioepiteliales gastrales. Recuerdan a las células mioepiteliales de la epidermis, pero son más altas y estrechas, con la parte contráctil menos desarrollada en la base, sin prolongaciones y con 2-5 flagelos en el lado apical que usa para mover los líquidos de la cavidad gastrovascular. Esta célula se caracteriza por la doble posibilidad de pinocitosis y fagocitosis. Recoge los materiales parcialmente digeridos de la cavidad gastrovascular y los pasan al resto de células. Otra característica es que su parte contráctil es perpendicular a la de la ectodermis.
- Células secretoras. Son células secretoras de tipo apocrino, altas, algunas flageladas, en cuyo interior se producen muchas sustancias (mucopolisacáridos, mucoproteínas, enzimas digestivas, etc.). Segregan sustancias que lubrifican y digieren los alimentos que entra en la cavidad gastrovascular.
Entre ellas, hay células sensoriales (en menor número que en la ectodermis) y células basales. También pueden aparecer cnidocitos. Plexo nervioso poco desarrollado.
Terminaciones nerviosas
editarEn algunos grupos aparecen receptores de diversa índole (táctiles, químicos, luminosos, gravitatorios, etc.)
Receptores táctiles
editarLos receptores táctiles son células ciliadas con prolongaciones típicas como las ya vistas en la anatomía interna.
Fotorreceptores
editar
Receptores para estímulos luminosos
Los más sencillos son las llamadas manchas ocelares, o manchas oculares, que son manchas pigmentadas que aparecen en ciertas medusas en la umbrela. Constituidas por grupos de células de dos tipos: sensoriales ciliadas y células en cuyo interior se acumula un pigmento (rodoxina). El pigmento se sitúa en la base del cilio de forma simétrica o asimétrica, en un replegamiento de membrana. La célula pigmentada envía información a la sensorial típica.
Más avanzados son los ocelos, que son un pequeño entrante de la epidermis en el cual las células fotorreceptoras quedan en el centro, rodeadas de células pigmentadas.
Este ocelo se complica hacia un ocelo en copa, en el que hay un entrante grande, en cuyo fondo las fotorreceptoras y rodeando las pigmentadas.
Un poco más allá, en algunas medusas se desprende la parte distal de la célula y se forman prolongaciones vacuolares que rellenan la hendidura, la "copa", formando una especie de lente.
Estos receptores no les permiten distinguir objetos. solo luces y sombras. Esto es lo que llamamos encelofalitis.
Órganos del equilibrio
editar
También han evolucionado diversos órganos del equilibrio.
Los estatos son estructuras que presentan filas de dos tipos de células, unas sensoriales ciliadas, y otras (litocitos) que acumulan en su interior una bolita calcárea (estatolito). Si el animal gira, como la célula con el estatolito cuelga por gravedad y pesa, se mueve y toca una célula sensorial de las que la rodea. Así el animal se mantiene informado de su posición.
Derivado de esto, tenemos los estatocistos, con una hendidura mayor que puede estar incluso cerrada y no comunicar con la umbrela. En otros casos, los estatolitos se desprenden de los litocitos o se usan partículas extrañas para la misma función.
También existen los estatorabdos, que son pequeños tentáculos con uno o más estatolitos. Este tentáculo cuelga rodeado de células sensoriales.
Ropalias
editarEn el orden escifozoos hay un órgano muy desarrollado, la ropalia, que es un centro quimio-estato-fotorreceptor. Es la estructura sensorial más compleja de los cnidarios, y solo aparece en la fase móvil. La ropalia más compleja de todas se encuentra en el orden cubozoos.
Toxinas en una picadura
editarLa estimulación mecánica y química de las barbas que rodean al cnido hace que una señal bioeléctrica mediada por calcio permita la salida del nematocisto a la superficie para expresar el veneno. La salida del nematocisto ocurre en 3 milisegundos y penetra hasta una profundidad de 0,9 mm, depositando la toxina a la microvasculatura de la dermis, para luego ser absorbida a la circulación sistémica mientras se ancla a la víctima.
La fracción tóxica del líquido contenido en los cnidocitos es una mezcla compleja de sustancias de naturaleza química, tales como catecolaminas, aminas bioactivas (histamina, serotoninas) bradicininas, colagenasas, hialuronidasas, proteasas, fosfolipasas, fibrinolisinas, dermatoneurotoxinas, cardiotoxinas, miotoxinas, nefrotoxinas, neurotoxinas y antígenos proteicos. El componente proteico de la toxina tiende a ser termolábil, no dializable, y se degrada por agentes proteolíticos. Además de las proteínas, también se han encontrado varias enzimas, aminas, hidratos de carbono y lípidos. Todas estas sustancias pueden tener efectos neurotóxicos, citotóxicos, a veces hemolíticos y cardiotóxicos. Las toxinas varían en función de los distintos tipos de cnidocitos y dentro de éstos también pueden diferir según las especies, pues existen venenos muy ligeros y también otros como el de la avispa de mar, que pueden llegar a causar la muerte.[6]
La toxina causa alteraciones en el transporte de los canales de sodio y de calcio, fracciona membranas celulares, libera mediadores inflamatorios y actúa como toxina directa en el miocardio, tejido nervioso, hepático y renal. También puede producir fuertes picores en el área renal de la persona.
Una vez se ha producido la picadura, hay que procurar no rascarse ni frotar sobre la zona en la que se nota la sensación de quemadura o el dolor intenso. No hay que lavarse con agua dulce, ya que el cambio osmótico hace que se disparen más cnidocistos, pero sí con agua salada. Hay que aplicar lo antes posible compresas frías durante 5-15 min. Se pueden preparar estas compresas con una bolsa de plástico llena de hielo, y no se debe aplicar la pieza de hielo directamente sobre la piel, ya que haría el mismo efecto que el agua dulce. La aplicación de compresas calientes está contraindicada, ya que el calor favorece la absorción sistémica del veneno.[7]
Función de algunas toxinas
editarLos venenos se van a dividir en 4 grupos, los cuales son enzimas, neurotoxinas, citolisinas y componentes bioactivos no proteicos. Las enzimas funcionarán como un virus, atacando a las células, infectándolas para así producir muchas más enzimas que ataquen al resto. Las citolisinas funcionaran muy parecido a las enzimas, la única diferencia es que estos atacarán en partes más específicas del cuerpo de la víctima, como alguno de los sistemas. Las neurotoxinas son capaces de atacar al sistema nervioso central y periférico principalmente, este daño provocará una serie de daños a los sistemas, además que muchas pueden ser degenerativas en la parte donde se llevó a cabo la picadura.[3] Para finalizar, los componentes bioactivos serán compuestos un poco más dolorosos en la zona de su picadura o que potencien el veneno, a pesar de ello, llegan a ser un poco menos agresivos para el resto de las sustancias tóxicas.
El veneno de estos cnidarios tendrá diferentes tipos de toxinas con distintos efectos, como
- Palitoxina: es un vasoconstrictor intenso y se considera una de las sustancias no proteicas más venenosas conocidas. En la imagen de al lado observamos la estructura de esta toxina.[8][9]

- Hipnocina: provoca somnolencia, parálisis muscular y respiratoria y tiene propiedades anestésicas.
- Thalassina: produce urticaria y edema en dosis pequeñas. A dosis elevadas puede causar parálisis nerviosa, paro cardiaco e incluso la muerte.
- Congestina: provoca congestión del tubo digestivo, vómitos, dolores abdominales, disminución de la presión sanguínea y problemas respiratorios.
- Tetramina: posiblemente con acción paralizante.
- 5 hidroxitriptamina: responsable del dolor y de la liberación de histamina.
- Actinoporinas: Familia multigénica de proteínas tóxicas producidas por diversas especies de anémonas. Estas proteínas actúan haciendo poros en las membranas plasmáticas de las células. En el esquema 3 se puede observar una representación de un poro formado por actinoporinas de Actinia fragacea sp.[10]

Ciclo
editar
El ciclo varía mucho según los grupos, por lo que para tener más información véanse las distintas clases. Aunque se caracteriza por la alternancia de generaciones.
De manera general, el huevo posee poco vitelo y sufre segmentación total e igual. En muchos la gastrulación es por delaminación para alcanzar un estado larvario general, la larva plánula, ciliada, nadadora, que buscará en el sustrato un lugar para fijarse, dando lugar al pólipo, que crece y en un momento determinado, por reproducción agamética (normalmente gemación)por un proceso denominado estrobilación, origina las medusas, en las que madurarán los gametos que formarán el nuevo huevo.
Este ciclo completo es un ciclo metagenético. Dentro de los grupos hay desviaciones; por ejemplo hay pólipos que dan pólipos (Hydra por gemación crea un pólipo que crece y se separa). También hay medusas cuyo huevo se desarrolla a plánula, pero que se desarrollará muy rápido a medusa. También hay fases intermedias; los sifonóforos son coloniales, y hay polipoides (derivados de pólipo) y medusoides (derivados de medusa) que coexisten en la colonia. Todos estos ciclos que no son completos se llaman hipogenéticos.
Clasificación
editarLa clasificación tradicional de los cnidarios reconoce cuatro clases y supone que los antozoos son la clase más primitiva y de la que han derivado las demás:
Solo pólipos (antopólipos), conocidos como anémonas y corales. No hay forma medusa. El pólipo da, por reproducción asexual o sexual, pólipos. Hay cnidocitos en la cavidad gastrovascular, a veces muy potentes, incluso en filamentos que salen por la pared del cuerpo. Las gónadas endodérmicas. La cavidad gastrovascular está dividida de forma completa (los tabiques provienen de la gastrodermis y de la mesoglea). El lado oral se introduce en la cavidad gastrovascular y origina un estomodeo o faringe.

Poseen exclusivamente medusas (cubomedusas), de forma cúbica, con cnidocitos especiales, muy potentes. Hasta hace poco se le consideraba como un orden de los escifozoos. Se caracterizan por su división tetrámera, que separa la cavidad gastrovascular en cuatro bolsas. El borde umbrelar no es festoneado y el margen de la subumbrela se pliega al interior para formar un velario.
Tienen hidropólipos e hidromedusas. Las medusas tienen velo (son medusas craspédotas). No tienen cnidocitos en la cavidad gastrovascular. Las gónadas son siempre de origen ectodérmico. En una sección transversal, la cavidad gastrovascular es sencilla, sin dividir. La mesoglea de los pólipos está poco desarrollada.
Posee pólipo pequeño e inconspicuo (escifopólipo o escifostoma) y medusa (escifomedusa), que carece de velo (acraspedota). Presentan cnidocitos en la cavidad gastrovascular. Las gónadas son endodérmicas. Una vez maduradas, de todas maneras, pueden almacenarse en la endodermis. Su cavidad gastrovascular está dividida incompletamente por 4 tabiques o septos incompletos en posición inter-radial (ver simetría radial); los tabiques separan 4 bolsas gastrales.
Actualmente, el Registro Mundial de Especies Marinas incluye también las siguientes clases en Cnidaria:
Son una clase de animales parásitos microscópicos, clasificados durante mucho tiempo como protozoos dentro de los esporozoos. La parasitación ocurre por esporas, con válvulas que contendrían uno o dos esporoblastos y una o dos cápsulas con filamentos que anclarían la espora al hospedador. En 2015 un estudio reveló que los mixozoos son en realidad cnidarios extremadamente reducidos en tamaños y con un genoma simplificado.[11]
Polipodiozoos (Polypodiozoa)
editarEs una clase monoespecífica de cnidarios parásitos. Es uno de los pocos cnidarios que viven en el interior de las células de otros animales. Los datos moleculares sugieren que podría estar relacionado con los mixozoos (también parásitos), pero este punto es controvertido, por lo que su clasificación es temporal, ya que sus relaciones filogenéticas no están claras.
Se trata de unas formas sésiles. La superficie aboral, correspondiente a la exumbrela de otras medusas, se prolonga en un tallo gracias al cual se fijan al sustrato, en especial algas y rocas. Hasta hace poco se consideraban un orden dentro de la clase Scyphozoa, pero se elevaron a la categoría de clase tras un estudio cladístico.[12]
Filogenia
editarSe han propuesto muchas teorías filogenéticas para relacionar a los distintos grupos de cnidarios. La mayor parte de los autores defienden que el posible pre-cnidario era un organismo planuloide adaptado a la vida sésil, adquiriendo simetría radial, que habría sufrido una invaginación que posteriormente dará a la cavidad gastrovascular. El pre-cnidario ¿tenía forma medusa o pólipo? Existen 2 teorías:[cita requerida]
| Teoría polipoide
Defiende que el pre-cnidario fue un pólipo
|
1 Simetría radial, cnidocitos, cavidad gastrovascular con tabiques, gónadas endodérmicas, tentáculos alrededor de la boca, cuerpo de forma polipoide, ADN mitocondrial circular; 2 cnidocitos con cnidocilo, ADN mitocondrial lineal; 3 medusa acraspedota; 4 ropalias; 5 ciclo metagenético con reducción de la fase pólipo; 6 estrobilación; 7 umbrela cúbica; 8 velario; 9 medusa craspedota; 10 pérdida de la tabicación de la cavidad gastrovascular; 11 adelgazamiento de la mesoglea y reducción de la cantidad de amebocitos; 12 gónadas ectodérmicas. | |||||||||||||||||||||||||||
| Teoría medusoide
Defiende que el pre-cnidario fue una medusa
|
1 Simetría radial, cavidad gastrovascular sin tabicar, gónadas ectodérmicas, tentáculos aldedor de la boca, larva plánula que se transforma en larva actínula; 2 pólipo, ciclo metagenético; 3 gónadas endodérmicas; 4 cavidad gastrovascular con tabiques; 5 reducción o pérdida de la fase pólipo; 6 ropalias; 7 estrobilación; 8 umbrela cúbica, velario; 9 pérdida de la fase medusa; 10 simetría hexarradial u octorradial; 11 faringe; 12 sifonoglifos; 13 cnidocitos sin cnidocilo ni opérculo. |
Estudios basados en filogenia molecular[13] corroboran la monofilia de los cnidarios y que la forma pólipo probablemente precedió a la forma medusa en la evolución de los cnidarios. También sugiere que los cnidarios están formados por dos grupos que podrían tener categoría de subfilos, los antozoos y los medusozoos; estos últimos agrupan los cubozoos, los escifozoos, los hidrozoos y las estauromedusas, por lo que las relaciones genómicas más recientes dan el siguiente resultado:[14][15][16]
| Cnidaria |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||








Referencias
editar- ↑ a b Fautin, Daphne Gail (15 de diciembre de 2009). «Structural diversity, systematics, and evolution of cnidae». Toxicon. Cnidarian Toxins and Venoms (en inglés) 54 (8): 1054-1064. ISSN 0041-0101. doi:10.1016/j.toxicon.2009.02.024. Consultado el 1 de febrero de 2021.
- ↑ Park, E.; Hwang, D.-S; Lee, J.-S; Song, J.-I; Seo, T.-K; Won, Y.-J (2012). «Estimation of divergence times in cnidarian evolution based on mitochondrial protein-coding genes and the fossil record.». Molecular Phylogenetics & Evolution 62 (1): 329-345. PMID 22040765. doi:10.1016/j.ympev.2011.10.008. Consultado el 30 de marzo de 2017.
- ↑ a b «Anémonas, corales y medusas: los cnidarios y su importancia médica». RDU UNAM. 24 de febrero de 2020. Consultado el 1 de febrero de 2021.
- ↑ Brusca, R. C. & Brusca, G. J., 2005. Invertebrados, 2ª edición. McGraw-Hill-Interamericana, Madrid (etc.), XXVI+1005 pp. ISBN 0-87893-097-3.
- ↑ Shadid, A.C., Shadid, O.B. (2002) GLOSARIO TETRALINGÜE DE TÉRMINOS APLICADOS A LA MORFOLOGÍA Y ANATOMÍA DE OCTOCORALLIA (COELENTERATA: ANTHOZOA). Filología y Lingüística XXVIII (2): 139-153.
- ↑ Jouiaei, Mahdokht; Yanagihara, Angel A.; Madio, Bruno; Nevalainen, Timo J.; Alewood, Paul F.; Fry, Bryan G. (2015/6). «Ancient Venom Systems: A Review on Cnidaria Toxins». Toxins (en inglés) 7 (6): 2251-2271. doi:10.3390/toxins7062251. Consultado el 1 de febrero de 2021.
- ↑ «Ciencia Canaria - Así pican las medusas en realidad». www.cienciacanaria.es. Consultado el 1 de febrero de 2021.
- ↑ Lazcano Pérez, José Fernando; Salazar Campos, Zayil; González-Márquez, Humberto (20 de marzo de 2020). «Anémonas, corales y medusas: los cnidarios y su importancia médica». Revista Digital Universitaria 21 (2). ISSN 1607-6079. doi:10.22201/codeic.16076079e.2020.v21n2.a4. Consultado el 1 de febrero de 2021.
- ↑ Jouiaei, Mahdokht; Yanagihara, Angel; Madio, Bruno; Nevalainen, Timo; Alewood, Paul; Fry, Bryan (18 de junio de 2015). «Ancient Venom Systems: A Review on Cnidaria Toxins». Toxins 7 (6): 2251-2271. ISSN 2072-6651. doi:10.3390/toxins7062251. Consultado el 1 de febrero de 2021.
- ↑ «SEBBM - Sociedad Española de Bioquímica y Biología Molecular.». www.sebbm.es. Consultado el 1 de febrero de 2021.
- ↑ E. Sally Chang, Moran Neuhof, Nimrod D. Rubinstein, Arik Diamant, Hervé Philippe, Dorothée Huchon, and Paulyn Cartwright. (2015) Genomic insights into the evolutionary origin of Myxozoa within Cnidaria. PNAS 2015, vol. 112 (48) 14912-14917; doi:10.1073/pnas.1511468112.
- ↑ Antonio C. Marques & Allen G. Collins. (2004). Cladistic analysis of Medusozoa and cnidarian evolution. Invertebrate Biology 123(1): 23-42.
- ↑ Collins, A. G., 2002. Phylogeny of Medusozoa and the evolution of cnidarian life cycles. Journal of Evolutionary Biology, 15(3):418-432.
- ↑ E. Sally Chang et al. 2015, Genomic insights into the evolutionary origin of Myxozoa within Cnidaria PNAS 2015 December, 112 (48) 14912-14917. https://doi.org/10.1073/pnas.1511468112
- ↑ Ou, Qiang; Han, Jian; Zhang, Zhifei; Shu, Degan; Sun, Ge; Mayer, Georg (31 de julio de 2017). «Three Cambrian fossils assembled into an extinct body plan of cnidarian affinity». Proceedings of the National Academy of Sciences 114 (33): 8835-8840. PMC 5565419. PMID 28760981. doi:10.1073/pnas.1701650114.
- ↑ Dunn, F. S.; Kenchington, C. G.; Parry, L. A.; Clark, J. W.; Kendall, R. S.; Wilby, P. R. (25 de julio de 2022). «A crown-group cnidarian from the Ediacaran of Charnwood Forest, UK». Nature Ecology & Evolution. doi:10.1038/s41559-022-01807-x.
[1]
Enlaces externos
editar Wikimedia Commons alberga una categoría multimedia sobre Cnidaria.
Wikimedia Commons alberga una categoría multimedia sobre Cnidaria. Wikispecies tiene un artículo sobre Cnidaria.
Wikispecies tiene un artículo sobre Cnidaria.
- ↑ Vera K, Cristián; Kolbach R, Marianne; Zegpi T, María Soledad; Vera K, Francisco; Lonza J, Juan Pedro (2004-02). «Picaduras de medusas:actualización: An update». Revista médica de Chile 132 (2). ISSN 0034-9887. doi:10.4067/s0034-98872004000200014. Consultado el 1 de febrero de 2021.