Heterokonta
Los heterocontos (Heterokonta) o estramenopilos (Stramenopiles) son una de las líneas evolutivas principales de Eukarya,[1] con unas 25 000 especies descritas.[3] El término "heteroconto" hace referencia a la presencia de flagelos desiguales, característica del grupo. El grupo incluye a organismos muy diversos, desde algas unicelulares como las diatomeas, que son componentes primarios del plancton, hasta las pluricelulares algas pardas, entre las que se encuentra el kelp, un tipo de alga gigante que forma bosques submarinos. Otros miembros notables son los oomicetos (generalmente parásitos), que superficialmente parecen hongos y que incluyen a Phytophthora, responsable de la plaga que asoló los cultivos de patata en Irlanda en el siglo XIX produciendo una gran hambruna, y Pythium que ocasiona la podredumbre de las semillas.[4]
| Heterokonta | ||
|---|---|---|
 Alga Ochromonas mostrando flagelos desiguales (heterocontos) | ||
| Taxonomía | ||
| Dominio: | Eukarya | |
| Reino: | Protista | |
| (sin rango) | Supergrupo SAR | |
| Superfilo: |
Heterokonta LÜTHER 1899 EM. CAVAL.-SM. 1981 o Stramenopiles PATTERSON 1989 | |
| Filos y clases[1][2] | ||
| ||
| Sinonimia | ||
| ||
Cavalier-Smith clasifica Heterokonta en cuatro grupos: Ochrophyta para las algas, Pseudofungi para los oomicetos, Bigyra conformado por fagótrofos no-fotosintéticos ciliados, flagelados y mohos mucilaginosos y Platysulcea como un clado ancestral a los anteriores.[5][6]
Características
editarLos heterocontos constituyen un grupo extenso y diverso que comprende tanto organismos fotosintéticos (algas) como fagotrofos (depredadores) y osmotrofos (comensales, parásitos y fungoides). Incluye desde organismos unicelulares y coloniales hasta verdaderos pluricelulares con diferenciación celular, como en el caso de las algas pardas. Incluye organismos marinos, de agua dulce, de suelo o parásitos de plantas y animales. La característica distintiva del grupo es la presencia en las etapas flageladas de dos flagelos heterocontos (desiguales), uno anterior mastigonemado y otro posterior liso.
Las células también pueden ser aflageladas, ciliadas, ameboides o formar parte de colonias, micelios (seudohongos), talos o verdaderos tejidos (algas pardas). Las células pueden estar desnudas, rodeadas de paredes celulares orgánicas o inorgánicas, o cubiertas de escamas y espinas. Los cloroplastos, cuando están presentes, están rodeados por cuatro membranas y presentan clorofilas a, c1 y c2, pues la clorofila b nunca está presente. Esto sugiere que los cloroplastos proceden de la endosimbiosis secundaria de un alga roja. Algunas formas presentan mancha ocular, bastante similar a la que tienen los euglénidos, y que utilizan en la fototaxis. Las mitocondrias tienen crestas tubulares.
Flagelos
editar
Muchos heterocontos son unicelulares flagelados y la mayoría de los demás producen células flageladas en algún punto de su ciclo vital, por ejemplo como gametos o zoosporas. El término heteroconto se refiere a la forma característica de estas células, que tienen típicamente dos flagelos desiguales. El flagelo anterior está cubierto con cerdas laterales o mastigonemas, mientras que el otro flagelo es liso y generalmente más corto o reducido a veces al cuerpo basal. Las cerdas del flagelo mastigonemado son tubulares tripartitas, esto es, presentan tres regiones. En algunas formas las cerdas están ausentes mientras que en otra se localizan sobre superficie del cuerpo. Los flagelos se insertan subapicalmente o lateralmente y se apoyan generalmente en cuatro raíces microtubulares con un patrón distintivo. Algunos grupos han perdido los flagelos.
Las mastigonemas están compuestas de glicoproteínas y son elaboradas en el retículo endoplasmático de la célula antes de ser transportadas a la superficie. Cuando el flagelo anterior se mueve, las mastigonemas crean una corriente contraria que tira de la célula a través del agua o le trae el alimento. Las mastigonemas tienen una estructura tripartita peculiar que se puede tomar como la característica que define al grupo, de tal modo se incluyen en el grupo algunos protistas que no producen células con la forma típica heteroconta. No obstante, las mastigonemas se han perdido en algunas líneas, siendo la de las diatomeas la más notable.
Cloroplastos
editar
Las algas heterocontas tienen cloroplastos rodeados por cuatro membranas, las dos primeras rodeando directamente al cloroplasto y las otras dos al retículo endoplasmático cloroplástido. La última de estas membranas es continua con el retículo endoplasmático y con la envoltura nuclear. Esta disposición de membranas sugiere que los cloroplastos de los heterocontos fueron adquiridos de un eucarionte simbiótico, probablemente de un alga roja. Los cloroplastos contienen típicamente clorofilas a, c1 y c2 y generalmente el pigmento asociado fucoxantina que les da un color marrón-amarillento o verde-terroso. Nunca presentan clorofila b ni nucleomorfo. Dentro de los cloroplastos los tilacoides se agrupan se apilan de tres en tres (estas pilas se denominan lamelas). Una de las lamelas se localiza alrededor de la periferia del cloroplasto, paralela y justo debajo de la envoltura del cloroplasto, a esta se le denomina lamela ceñidora.
La mayoría de los heterocontos basales no están coloreados, lo que parece sugerir que la separación tuvo lugar antes de la aparición de cloroplastos en el grupo. Sin embargo, también se encuentran cloroplastos conteniendo fucoxantina en Haptophyta por lo que es posible que ambos grupos compartan un origen común. En vista de esto, es posible que el heteroconto ancestral fuera un alga y todos los grupos no coloreados sufrieron posteriormente la pérdida de cloroplastos.
Galería
editar-
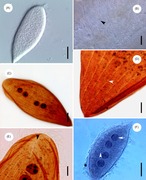
-

-
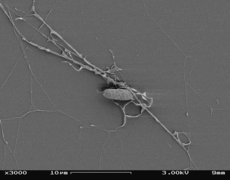
-
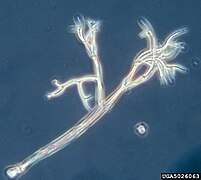
-

-

-

-
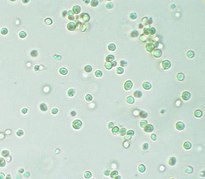
-
-
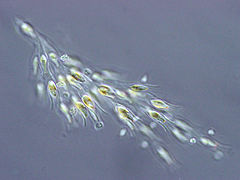
 Colonia de Synura (Synurophyceae)
Colonia de Synura (Synurophyceae) -
-

-

-















Filogenia
editarLos heterocontos son excepcionalmente diversos y su clasificación se realiza casi exclusivamente sobre la base de análisis moleculares, pues la morfología de los distintos grupos es tan diversa que hace imposible establecer caracteres homólogos. Los análisis moleculares respaldan que los heterocontos comprenden tres líneas de organismos: los heterótrofos Platysulcea, Bigyra, Pseudofungi y los predominantemente fotosintéticos Ochrophyta. La separación más temprana tuvo lugar entre Platysulcea y las restantes dos líneas. No está claro si el heteroconto ancestral era heterótrofo y los cloroplastos fueron obtenidos posteriormente por Ochrophyta, o por el contrario, el heteroconto ancestral era fotosintético y posteriormente las otras tres líneas sufrieron la pérdida de cloroplastos.[7]
El siguiente árbol filogenético es un consenso de los publicados por varios autores:[7][8][9][10][11]
| Heterokonta |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Platysulcea
editarContiene el género Platysulcus el cual es un protozoo biflagelado ovoide fagotrófico de vida libre, bacterívoro y marino relacionado con el antepasdo común de los heterocontos.
Bigyra
editarEl antepasado de Bigyra y probablemente de todos los heterocontos era un organismo fagotrofo de vida libre. Bikosea, Placididea son los únicos grupos de esta línea que retienen el modo de alimentación ancestral fagotrófica. Posteriormente, Opalinea y Blastocystea reemplazaron la capacidad fagotrófica ancestral por una osmotrofia secundaria cuando se convirtieron en comensales o parásitos de los animales. Por su parte, Labyrinthulea probablemente perdió su capacidad fagotrófica independientemente cuando se adaptó a un modo de vida saprobio de tipo fungoide.
Pseudofungi
editarLa línea Pseudofungi comprende organismos con un tipo de alimentación osmotrofa, aunque como excepción, un par de géneros retienen el modo de alimentación fagotrófica. Los antepasados de Oomycetes probablemente surgieron en el mar parasitando las algas heterocontas. El antepasado de Hyphochytridiomycetes fue probablemente un depredador que utilizaba sus seudópodos para penetrar a sus presas.
Ochrophyta
editarLa clasificación de los heterocontos fotosintéticos o Ochrophyta también se realiza sobre la base análisis moleculares, pues no es posible comparar directamente la pared celular de celulosa de las algas pardas, la frústula silícea de las diatomeas y las células desnudas de las algas verde-amarillas. Así, las algas heterocontas comprenden cinco clados principales, obtenidos mediante análisis filogenéticos, que a su vez se pueden agrupar en tres grupos:[12]
- Grupo III o Diatomista: líneas Khakista (clases Bolidophyceae y Bacillariophyceae) e Hypogyrista (clases Dictyochophyceae y Pelagophyceae).
- Grupo II o Limnista: líneas Eustigmista (clases Pinguiophyceae y Eustigmatophyceae) y Phagochrysia (clases Picophagophyceae, Chrysophyceae y Synurophyceae).
- Grupo I o Marista: clases Raphidophyceae, Schizocladiophyceae, Xanthophyceae, Phaeothamniophyceae, Phaeophyceae, además de Actinophryida, que son organismos heterótrofos fagotrofos derivados de Raphidophyceae.
La composición de los carotenos de las algas heterocontas puede dar una idea de su pasado evolutivo. La mayoría de los grupos de protistas con clorofila c realizan el ciclo diatoxantina-diadinoxantina (ciclo D-D), que puede asumirse como el ciclo original de los heterocontos ancestrales. El grupo III presenta solo este ciclo, por lo que sería la línea basal de las algas heterocontas. El grupo II, por el contrario, presenta solo el ciclo violaxantina-anteraxantina (ciclo V-A), mientras que en el grupo I existen grupos con ambos ciclos. Esto es, probablemente el ciclo V-A apareció por primera vez en el grupo I y luego fue heredado por sus descendientes del grupo II.
Los miembros del grupo III tienden a la reducción del aparato flagelar, por ejemplo, carecen de raíces flagelares microtubulares y las células nadadoras tienen un solo flagelo emergente. Los centrosomas se unen directamente al núcleo celular que está desplazado anteriormente y ancla el flagelo al cuerpo celular. Esta reducción flagelar fue llevada al extremo en las diatomeas pennadas, en las que en la actualidad no se aprecian restos de flagelos.
Referencias
editar- ↑ a b Adl et al. 2012. The revised classification of eukaryotes. Journal of Eukaryotic Microbiology, 59(5), 429-514
- ↑ Silar, P. (2016). Protistes Eucaryotes.
- ↑ Chapman, A. D. (2009). Numbers of living species in Australia and the world Archivado el 28 de septiembre de 2015 en Wayback Machine..
- ↑ Blackwell, W. H. (2009). «Chromista revisited: A dilemma of overlapping putative kingdoms, and the attempted application of the botanical code of nomenclature» (PDF). Phytologia 91 (2).
- ↑ Cavalier-Smith, T., & Chao, E. E. (2006). Phylogeny and megasystematics of phagotrophic heterokonts (kingdom Chromista). Journal of molecular evolution, 62(4), 388-420.
- ↑ Riisberg I, Orr RJ, Kluge R, etal (May 2009). «Seven gene phylogeny of heterokonts». Protist 160 (2): 191-204. PMID 19213601. doi:10.1016/j.protis.2008.11.004.
- ↑ a b Cavalier-Smith, Thomas, and Josephine Margaret Scoble. Phylogeny of Heterokonta: Incisomonas marina, a uniciliate gliding opalozoan related to Solenicola (Nanomonadea), and evidence that Actinophryida evolved from raphidophytes. European journal of protistology 49.3 (2013): 328-353.
- ↑ Shiratori, Takashi; Nakayama, Takeshi; Ishida, Ken-ichiro (2015). «A New Deep-branching Stramenopile, Platysulcus tardus gen. nov., sp. nov.». Protist 166 (3): 337-348. ISSN 1434-4610. PMID 26070192. doi:10.1016/j.protis.2015.05.001. hdl:2241/00148461.
- ↑ Anna Cho, Denis V. Tikhonenkov, Elisabeth Hehenberger, Anna Karnkowska, Alexander P. Mylnikov, Patrick J. Keeling (2021). Monophyly of Diverse Bigyromonadea and their Impact on Phylogenomic Relationships Within Stramenopiles. Biorxiv.
- ↑ Thakur R, Shiratori T, Ishida KI (November 2019). «Taxon-rich Multigene Phylogenetic Analyses Resolve the Phylogenetic Relationship Among Deep-branching Stramenopiles». Protist 170 (5): 125682. PMID 31568885. doi:10.1016/j.protis.2019.125682.
- ↑ Cavalier-Smith, Thomas; Scoble, Josephine Margaret (2013). «Phylogeny of Heterokonta: Incisomonas marina, a uniciliate gliding opalozoan related to Solenicola (Nanomonadea), and evidence that Actinophryida evolved from raphidophytes». European Journal of Protistology 49 (3): 328-353. PMID 23219323. doi:10.1016/j.ejop.2012.09.002.
- ↑ Yang, E. C., Boo, G. H., Kim, H. J., Cho, S. M., Boo, S. M., Andersen, R. A., & Yoon, H. S. (2012). Supermatrix data highlight the phylogenetic relationships of photosynthetic stramenopiles Archivado el 13 de abril de 2015 en Wayback Machine.. Protist, 163(2), 217-231.
Enlaces externos
editar Wikimedia Commons alberga una categoría multimedia sobre Heterokonta.
Wikimedia Commons alberga una categoría multimedia sobre Heterokonta.- Árbol de la vida del Proyecto Vida Web: Stramenopiles Archivado el 19 de octubre de 2003 en Wayback Machine.
- Universidad de Penn State: Stramenopila (también los Rhodophyta, Chlorophyta)
- Phylogeny and Megasystematics of Phagotrophic Heterokonts (Kingdom Chromista) (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última).
 Datos: Q461622
Datos: Q461622- Multimedia: Stramenopiles / Q461622
 Especies: Stramenopiles
Especies: Stramenopiles