Metamonada
Metamonada es un extenso grupo de protistas flagelados inusuales por carecer de mitocondrias.[2][1][3] Son todos anaerobios y en su mayoría simbiontes de animales. Ciertas especies viven en el tracto digestivo de las termitas, desempeñando un papel importante en la degradación de la celulosa. Algunas formas son parásitas, incluyendo algunas que parasitan a los seres humanos, como Trichomonas y Giardia. Los vestigios mitocondriales incluyen mitosomas e hidrogenosomas, que producen el hidrógeno. Las células pueden tener uno o dos núcleos. Todos los subgrupos comparten el hecho de tener flagelos en grupos característicos de cuatro, que a menudo se asocian a los núcleos. El grupo comprende unas 710 especies conocidas.[4]
| Metamonada | ||
|---|---|---|
 | ||
| Taxonomía | ||
| Dominio: | Eukaryota | |
| Reino: | Protista | |
| (sin rango) | Opimoda ? | |
| Filo: |
Metamonada Grassé 1952 emend. | |
| Clases y órdenes[1] | ||
Retortamonadida Trichomonadida
| ||
Morfología
editarMetamonada comprende organismos anaerobios que abundan en hábitats acuáticos pobres en oxígeno alimentándose de bacterias mediante fagocitosis. También abundan y se alimentan de manera similar en el tracto digestivo (pobre en oxígeno) de muchos animales. Así, se encuentran como simbiontes en el intestino de las termitas y de otros invertebrados, a los que ayudan a digerir la celulosa. Sin embargo, otros son comensales o parásitos, capaces de producir enfermedades en los seres humanos.[5]
Son organismos flagelados fagotrofos que se caracterizan por presentar, al menos ancestralmente, pues se ha perdido secundariamente en algunos grupos, una boca o surco de alimentación ventral soportada por microtúbulos, típica de los excavados. Así, los miembros de Fornicata suelen presentar una ranura de alimentación o tubo citofaringeal asociado a cada cinétida. En Parabasalia, el surco de alimentación está ausente, pero se supone que es una pérdida secundaria. Por último, en Preaxostyla la ranura falta en Oxymonadida, aunque se presenta en Trimastix, un organismo íntimamente relacionado con los anteriores.
La característica distintiva del grupo y que los hace inusuales es la carencia de mitocondrias. En el pasado eran considerados los Eukarya más primitivos, divergiendo del resto antes de que aparecieran las mitocondrias. Sin embargo, ahora se sabe que han perdido las mitocondrias secundariamente y que conservan los orgánulos y genes nucleares derivados de ellos. Los vestigios mitocondriales incluyen los hidrogenosomas, que producen el hidrógeno, las pequeñas estructuras llamadas mitosomas y, en algunas especies, también se han descubierto genes mitocondriales en el genoma nuclear.
Los flagelos están dispuestos en uno o más grupos cerca de la parte anterior de la célula, usualmente en conjuntos característicos de cuatro. Probablemente el antecesor de estos organismos tenía cuatro flagelos y cuatro centriolos. Aunque algunas especies tienen solo dos flagelos y otras tienen cientos de ellos. Los flagelos a menudo se asocian al núcleo, formando una estructura llamada cariomastigonte (de cario, núcleo y mastigonte, órgano de movilidad). Es decir, que el núcleo está conectado al aparato flagelar. Diplomonadida tiene una estructura celular doble, con componentes celulares duplicados, entre ellos dos núcleos y dos conjuntos de flagelos.
Muchos se caracterizan por presentar elaborados citoesqueletos de numerosas fibras distintivas de proteínas. Así, muchos presentan un axostilo, un conjunto de varios miles de microtúbulos que soportan la célula a modo de esqueleto. En Parabasalia el axostilo permanece inmóvil, mientras que en Oxymonadida se mueve libremente dentro de la célula. Parabasalia también se caracteriza por presentar un aparato parabasal, constituido por dos o más fibras parabasales estriadas que conectan el aparato de Golgi al sistema flagelar. Sin embargo, otros grupos como Oxymonadida carecen del aparato de Golgi.
Clasificación
editarMetamonada incluye tres grupos en su mayoría simbiontes, comensales o parásitos en el tracto digestivo de los animales, aunque también hay especies de vida libre que se alimentan de bacterias.[4]

Fornicata
editarLos miembros de Fornicata o Eopharyngia presentan o bien y un único núcleo celular y una sola cinétida o bien un par de núcleos y cinétidas. Además tienen dos cinetosomas por cinétida y, por lo general, una ranura de alimentación o tubo citofaringeal asociado a cada cinétida. Carecen de mitocondrias y de hidrogenosomas, aunque a veces presentan un orgánulo rodeado por membranas que parece ser los restos de un antiguo hidrogenosoma.
Son en su mayoría comensales o parásitos del tracto digestivo de animales, aunque hay especies de vida libre. Así, por ejemplo, Retortamonas intestinalis, Giardia lamblia, Chilomastix mesnili y Enteromonas hominis parasitan el tracto digestivo de humanos y otros primates.
Retortamonas (incluido en Retortamonadida) tiene un único núcleo, dos flagelos emergentes y un surco de alimentación ventral. Hexamita y Giardia (incluidos en Diplomonadida) presentan dos núcleos y ocho flagelos dispuestos en dos grupos iguales de cuatro flagelos.[6] Cada núcleo está asociado a un grupo flagelar y con un conjunto de cinetosomas asociadas. Hexamita tiene dos citostomas que se abren en la parte posterior de la célula. Giardia tiene un cuerpo aplanado ventralmente y dispone de un disco adhesivo soportado por un complejo citoesqueleto; no presenta citostomas; los dos grupos de flagelos están dispuestos juntos en el centro de la célula, entre los dos núcleos. Carpediemonas (incluido en Carpediemonadida) es un organismo de vida libre, considerado basal a los grupos anteriores.
Parabasalia
editarLos miembros de Parabasalia se caracterizan por presentar un aparato parabasal, constituido por dos o más fibras parabasales estriadas que conectan el aparato de Golgi al sistema flagelar. Las cinétidas generalmente presentan cuatro cinetosomas y flagelos, aunque en algunos grupos se produce la reducción o pérdida del aparato flagelar, mientras que en otros se produce la multiplicación de todos o parte de los elementos (algunas especies puede llegar tener hasta mil flagelos). Presentan hidrogenosomas en lugar de las mitocondrias. Carecen de los surcos de alimentación encontrados en los demás grupos, pero esto es probable que también sea una pérdida secundaria.
La mayoría son simbiontes o parásitos de animales, incluyendo una gran variedad de especies encontradas en el tubo digestivo de termitas y cucarachas, muchas de las cuales tienen bacterias simbióticas que les ayudan a digerir la madera. También incluye algunos patógenos para los seres humanos, como Trichomonas, Pentatrichomonas hominis y Dientamoeba fragilis.
Preaxostyla
editarLos miembros de Preaxostyla o Anaeromonadea presentan cuatro flagelos y cinetosomas por cinétida y carecen de las mitocondrias clásicas. Se caracterizan por la presencia de un preaxostilo, que es una fibra que conecta los centrosomas con el axostilo. Este último es un orgánulo formado por microtúbulos que recorre la célula axialmente, y que es de diferente estructura a los axostilos de los parabasálidos.
Oxymonadida comprende especies que viven exclusivamente en los intestinos de termitas y de otros insectos comedores de madera. De forma similar a los parabasálidos contienen bacterias simbióticas que les facilitan la digestión de la celulosa. Se caracterizan por carecer del aparato de Golgi. Trimastix es un organismo de vida libre relacionado con los anteriores, pero al contrario que estos presenta un aparato de Golgi típico.
Filogenia
editarSe ha postulado que las relaciones son las siguientesː[7]
| Metamonada |
| ||||||||||||
Especies notables
editar- Enteromonas hominis. Parasita el tracto digestivo de humanos y otros mamíferos.
- Chilomastix mesnili. Parasita el tracto digestivo de humanos y otros primates.
- Giardia lamblia. Parasita el tracto digestivo de humanos y otros mamíferos, produciendo una patología denominada giardiasis.
- Retortamonas intestinalis. Parasita el tracto digestivo de humanos y otros primates.
- Trichomonas vaginalis. Parasita el tracto urogenital tanto de hombres como de mujeres, pero únicamente en humanos. Produce una hemorragia denominada tricomoniasis urogenital.
- Trichomonas tenax. Parasita exclusivamente la cavidad bucal de los seres humanos.
- Pentatrichomonas hominis. Parasita principalmente el intestino grueso de los seres humanos y otros mamíferos.
- Dientamoeba fragilis. Parasita principalmente el colon humano.
Galería
editar-

-
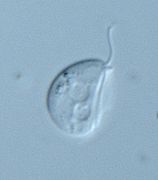
-
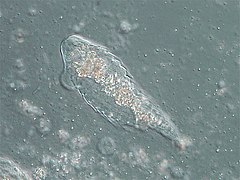 Hypermastigida (Parabasalia)
Hypermastigida (Parabasalia) -
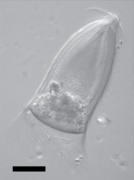
-
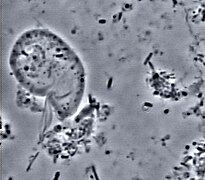





Véase también
editarReferencias
editar- ↑ a b Adl, S.M. et al. 2012. The revised classification of eukaryotes. Journal of Eukaryotic Microbiology, 59(5), 429-514
- ↑ T. Cavalier-Smith (2003). «The excavate protozoan phyla Metamonada Grassé emend. (Anaeromonadea, Parabasalia, Carpediemonas, Eopharyngia) and Loukozoa emend. (Jakobea, Malawimonas): their evolutionary affinities and new higher taxa». International Journal of Systematic and Evoluionary Microbiology 53: 1741-1758.
- ↑ Laura Wegener Parfrey, Erika Barbero, Elyse Lasser, Micah Dunthorn, Debashish Bhattacharya, David J Patterson, and Laura A Katz (2006 December). «Evaluating Support for the Current Classification of Eukaryotic Diversity». PLoS Genet. 2 (12): e220. doi:10.1371/journal.pgen.0020220.
- ↑ a b Ald, S.M. et al. (2007) Diversity, Nomenclature, and Taxonomy of Protists, Syst. Biol. 56(4), 684–689, DOI: 10.1080/10635150701494127.
- ↑ Cavalier-Smith, T. (2013). Early evolution of eukaryote feeding modes, cell structural diversity, and classification of the protozoan phyla Loukozoa, Sulcozoa, and Choanozoa. European journal of protistology, 49(2), 115-178.
- ↑ a b Jeffrey D. Silberman, Alastair G. B. Simpson, Jaroslav Kulda, Ivan Cepicka, Vladimir Hampl, Patricia J. Johnson and Andrew J. Roger, Retortamonad Flagellates are Closely Related to Diplomonads—Implications for the History of Mitochondrial Function in Eukaryote Evolution, Molecular Biology and Evolution, 19:777-786 (2002).
- ↑ T. Cavalier-Smith 2003, The excavate protozoan phyla Metamonada Grasse´ emend. (Anaeromonadea, Parabasalia, Carpediemonas, Eopharyngia) and Loukozoa emend. (Jakobea, Malawimonas): their evolutionary affinities and new higher taxa. International Journal of Systematic and Evolutionary Microbiology (2003), 53, 1741–1758 DOI 10.1099/ijs.0.02548-0
 Datos: Q499091
Datos: Q499091 Multimedia: Metamonada / Q499091
Multimedia: Metamonada / Q499091 Especies: Metamonada
Especies: Metamonada