Perissodactyla
Los perisodáctilos, Perissodactyla o ungulados de dedos impares (del griego antiguo περισσός perissós, "impar"; y δάκτυλος dáktylos, "dedo"), son un orden de mamíferos placentarios. Son mamíferos ungulados que se caracterizan por la posesión de extremidades con un número impar de dedos terminados en pezuñas, y con el dedo central, que sirve de apoyo, más desarrollado que los demás. Son herbívoros. En la actualidad solo incluye a los caballos, asnos, cebras (suborden Hippomorpha), los tapires y los rinocerontes (suborden Ceratomorpha).
| Perissodactyla | ||
|---|---|---|
| Rango temporal: 58,7 Ma - 0 Ma Paleoceno - Reciente[1] | ||
 | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Subfilo: | Vertebrata | |
| Clase: | Mammalia | |
| Subclase: | Theria | |
| Infraclase: | Placentalia | |
| Magnorden: | Boreoeutheria | |
| Superorden: | Laurasiatheria | |
| Granorden: | Scrotifera | |
| Orden: |
Perissodactyla Owen, 1848 | |
| Subórdenes | ||
Características
editar
Los primeros perisodáctilos eran muy similares a los otros ungulados primitivos. Las patas desarrollan la condición mesaxónica, es decir, el dedo central (tercero) se hace más grande y los demás se reducen, al mismo tiempo que la región inferior de las extremidades se alarga y la superior se acorta, con reducción de la ulna (cúbito) y la fíbula (peroné).
Un rasgo distintivo de los perisodáctilos es la organización del carpo y del tarso. En las patas anteriores (a veces referidas como "manos" en el habla popular), un carpiano distal, el hueso grande, se ensancha y se articula con los carpianos proximales, mientras que en el pie, el ectocuneiforme se transforma en un gran hueso plano que transmite el empuje del astrágalo a través de un navicular aplanado; el astrágalo tiene la superficie inferior aplanada y no a modo de tróclea como en los artiodáctilos.[2]
El aparato digestivo está menos especializado que en los artiodáctilos. Los incisivos se conservan y se usan para segar la hierba, los caninos están reducidos o ausentes y, a menudo, hay un diastema. Los molares de las formas primitivas permanecieron bunodontos y de corona baja, pero en los rinocerontes y caballos modernos tienen una elaborada superficie trituradora; los premolares se molarizan, con lo que se consigue un gran superficie trituradora.
El tubo digestivo es más simple que el de los artiodáctilos. El estómago no presenta cámaras. La digestión de la celulosa se realiza en el ciego y el intestino grueso, que puede estar muy desarrollado.
Los perisodáctilos son macrosmáticos, es decir, predomina el sentido del olfato, con la porción sensorial de la nariz muy desarrollada.
Anatomía interna
editarLa estructura del tubo digestivo de los ungulados pares difiere mucho de la de los ungulados pares, en su mayoría herbívoros. Al igual que los roedores, los ungulados pares son fermentadores rectales, lo que significa que la digestión tiene lugar principalmente en el intestino grueso. A diferencia de los ungulados pares, el estómago es siempre simple y unicameral; la fermentación tiene lugar en el recto, de gran tamaño (que puede contener 90 litros). (que puede contener hasta 90 litros en los caballos) y en el intestino de doble bucle. El intestino es muy largo (hasta 26 metros en los caballos). La utilización de los alimentos es relativamente baja, lo que probablemente ha provocado que hoy en día ya no existan ungulados pares de pequeño tamaño, ya que la necesidad de alimento por kilogramo de peso corporal es menor en los animales grandes y la relación superficie-volumen es menor (lo que es mejor para el balance térmico).
En el área del tracto urogenital, las hembras se caracterizan originalmente por un "útero bicorne" (útero bicorne). El ovarios (ovarios) en rinocerontes y tapires se encuentran en una bolsa del peritoneo (bolsa ovárica, bursa ovarica); en los caballos, la bolsa ovárica sólo cubre parcialmente el ovario. Los caballos difieren del resto de mamíferos en la estructura del ovario: En los caballos, el tejido ovárico denominado habitualmente "corteza", con los folículos, se encuentra en el interior del órgano, mientras que la médula ovárica vascularizada se encuentra en el exterior. La corteza ovárica sólo llega a la superficie en un punto. Este punto también es visible desde el exterior como una retracción y se conoce como "fosa ovárica" (Fossa ovarii); la ovulación sólo puede tener lugar en este punto. En los ungulados impares machos, los testículos de rinocerontes y tapires son inguinales (en la región inguinal), sólo los caballos tienen escroto.
Cráneo y dientes
editar
Los ungulados pares tienen una cabeza alargada, por lo que una larga mandíbula superior (maxilar superior). Las diferentes formas del hocico de las distintas familias se deben a diferencias en la estructura de la premaxila. El hueso lagrimal (lacrimal) tiene una joroba que sobresale en la cuenca ocular; una autapomorfía es el amplio contacto entre el hueso lagrimal y el hueso nasal (nasales). Otro rasgo característico es una mandíbula maciza, sobre todo en las especies herbívoras. La articulación temporomandibular es alta y la rama de la mandíbula inferior está agrandada.
Los rinocerontes tienen uno o dos cuernos que, a diferencia de los cuernos de los ungulados de dedos pares, no están formados por sustancia ósea, sino por queratina aglutinada.
El número y la estructura de los dientes varían según el alimento. Los incisivos y caninos pueden ser muy pequeños o estar completamente ausentes (por ejemplo, en las dos especies de rinocerontes africanos; en los caballos, normalmente sólo los machos tienen caninos - "diente ganchudo"). Debido a la mandíbula superior alargada, existe un hueco conocido como diastema entre los dientes frontales y los dientes mandibulares. Los premolares están en su mayoría están desarrollados al modo de molares; la forma de la superficie y la altura de los molares depende en gran medida de si el principal componente de la dieta es el follaje blando o la hierba dura. Hay tres o cuatro premolares y siempre tres molares por mitad de la mandíbula, de modo que la fórmula dentaria de los ungulados impares es: I 0-3/0-3 - C 0-1/0-1 - P 2-4/2-4 - M 3/3.
Extremidades
editarEl eje principal de las patas delanteras y traseras pasa por la ranilla central y, por tanto, el tercer dedo es el más grande de todas las especies. Los rayos restantes se han reducido en diversos grados, y menos aún en los tapires. Estos animales tienen cuatro dedos en las patas delanteras, una adaptación al suelo blando de su hábitat, y tres en las traseras. Los rinocerontes actuales tienen tres dedos en las patas delanteras y traseras. En los caballos, la reducción de los radios laterales es más avanzada; estos animales sólo tienen un dedo (monodactilia). Las patas tienen pezuñas, que en los caballos, sin embargo, sólo cubren los dedos casi por completo; En los rinocerontes y tapires, solo el borde frontal está cubierto con una pezuña, la parte inferior es blanda; los rinocerontes también tienen una almohadilla de suela blanda[3].
Dentro de las patas, el cúbito y el peroné son de tamaño reducido; en los caballos, estos huesos incluso están fusionados al radio y la tibia en la mitad inferior . Una autapomorfia (una característica común que distingue claramente a este grupo de otros grupos) es la articulación talonavicular en forma de silla de montar, la articulación del tobillo entre el hueso del tobillo (astrágalo) y el hueso escafoides (navicular), que limita gravemente la movilidad. El muslo es relativamente corto y falta la clavícula.
Carencia de rete carotídea
editarA diferencia de los artiodáctilos, los perisodáctilos carecen de rete carótida,[4] un intercambio de calor que reduce la dependencia de la temperatura del cerebro respecto a la del cuerpo. Como resultado, los perisodáctilos tienen una flexibilidad termorreguladora limitada en comparación con los artiodáctilos, lo que los ha restringido a hábitats de baja estacionalidad y ricos en alimento y agua, como los bosques tropicales. Por el contrario, los artiodáctilos ocupan una amplia gama de hábitos que van desde el círculo polar ártico hasta los desiertos y las sabanas tropicales.[5]
Reproducción y desarrollo
editarLos ungulados pares se caracterizan por un largo periodo de gestación y una camada de pequeño tamaño; por regla general, nace una sola cría. El periodo de gestación oscila entre 330 y 500 días, siendo el más largo el de los rinocerontes. Los ungulados impares recién nacidos son fugitivos del nido, los caballos jóvenes y los rinocerontes pueden seguir a su madre tras sólo unas horas; sólo las crías de tapir pasan sus primeros días de vida en un campamento protegido[3].
Las crías son amamantadas durante un tiempo relativamente largo, a menudo hasta su segundo año de vida, y alcanzan la madurez sexual en torno a los dos u ocho años de edad. Suelen ser animales longevos; se ha documentado que varias especies viven casi 50 años bajo cuidado humano.
Estilo de vida y alimentación
editarEn función de su hábitat, las distintas especies de ungulados impares llevan estilos de vida diferentes. Suelen ser animales crepusculares o nocturnos. Los tapires viven en solitario y habitan principalmente en selvas tropicales y otros bosques. Los rinocerontes también suelen vivir como animales solitarios y se encuentran en sabanas secas de África y en zonas húmedas de pantanos o bosques de Asia. Por último, los caballos habitan en zonas abiertas como praderas, estepas o semidesiertos y viven en grupo. Los ungulados pares son exclusivamente herbívoros que se alimentan en mayor o menor medida de hierbas, hojas y otras partes de las plantas[6]. Se suele distinguir entre las formas predominantemente herbívoras (rinoceronte blanco, solípedos) y las que se alimentan de hojas (tapires, otros rinocerontes).
Historia evolutiva
editarLos perisodáctilos más primitivos se habían separado muy poco de los condilartros. Hyracotherium, del Eoceno tenía el tamaño de un perro pequeño y se parecía al condilartro Phenacodus; tenía una dentición completa, con premolares trituberculados y molares cuadrados bunodontos con dos crestas transversales. La locomoción era digitígrada en las formas primitivas.
Los primeros perisodáctilos conocidos datan del Eoceno, aunque posiblemente surgieron en Asia durante el Paleoceno Superior, menos de 10 millones de años después de la extinción masiva del Cretácico-Terciario en la cual se extinguieron los dinosaurios y otros muchos organismos.[7] A principios de Eoceno, hace 55 millones de años, ya estaban diversificados y ocupaban varios continentes.
Los caballos y tapires evolucionaron en Norte América[8] mientras que los rinocerontes parecen haber evolucionado en Asia a partir de animales similares al tapir y luego haber recolonizado América durante el Eoceno Medio (hace unos 45 millones de años). Existieron 15 o 16 familias, de las cuales solo tres sobreviven en la actualidad.[7][9]
Dichas familias fueron muy diversas en apariencia y tamaño; algunas incluían animales gigantescos (Brontotheriidae) y extraños (Chalicotheriidae). El mayor perisodáctilo fue un rinoceronte asiático (Paraceratherium) que, con 11 toneladas, fue más de dos veces mayor que los elefantes actuales.
Los perisodáctilos fueron el grupo de grandes mamíferos herbívoros dominante durante el Oligoceno. No obstante, la expansión de las praderas durante el Mioceno (hace unos 20 millones de años) favorecieron a los artiodáctilos que, con sus estómagos provistos de cámaras se adaptaron mejor a una dieta tan pobre en nutrientes y pronto les arrebataron la supremacía. A pesar de ello, muchas especies de perisodáctilos sobrevivieron y prosperaron hasta el final del Pleistoceno (hace solo 10 000 años), cuando no pudieron soportar la presión de los cazadores humanos ni el nuevo hábitat.
-

-

-
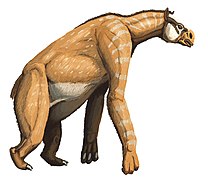 Un calicotérido
Un calicotérido -
 Esqueleto de brontoterio
Esqueleto de brontoterio




Taxonomía
editar
La siguiente clasificación (hasta el nivel de familia) está basada en el estudio de McKenna & Bell (1997)[9][10]
Orden Perissodactyla
- Suborden Hippomorpha
- Superfamilia Pachynolophoidea †
- Familia Pachynolophidae †
- Superfamilia Equoidea
- Familia Palaeotheriidae †
- Familia Indolophidae †
- Familia Equidae
- Superfamilia Pachynolophoidea †
- Suborden Ceratomorpha
- Clado Selenida
- Superfamilia Brontotheroidea †
- Familia Anchilophidae †
- Familia Lambdotheriidae †
- Familia Brontotheriidae †
- Superfamilia Chalicotheroidea †
- Familia Chalicotheriidae †
- Superfamilia Brontotheroidea †
- Clado Selenida
- Clado Tapiromorpha
- Superfamilia Rhinocerotoidea
- Familia Amynodontidae †
- Familia Hyrachyidae †
- Familia Hyracodontidae †
- Familia Rhinocerotidae
- Superfamilia Tapiroidea
- Familia Isectolophidae †
- Familia Helaletidae †
- Familia Lophiodontidae †
- Familia Deperetelidae †
- Género Bahinolophus †
- Género Deperetella †
- Familia Lophialetidae †
- Familia Tapiridae
- Superfamilia Rhinocerotoidea
- Clado Tapiromorpha

Cladograma según Hooker y Dashzeveg de 2004:[11]
| Perissodactyla |
| ||||||||||||||||||||||||||||||||||||||||||||||||
Referencias
editar- ↑ Perissodactyla PBDB
- ↑ Young, J. Z. 1977. La vida de los vertebrados. Editorial Omega, Barcelona, 660 pp. ISBN 84-282-0206-0
- ↑ a b Thomas S. Kemp: The Origin & Evolution of Mammals. Oxford University Press, Oxford 2005. ISBN 0-19-850761-5.
- ↑ Mitchell, G; Lust, A (23 de agosto de 2008). «La reta carótida y el éxito de los artiodáctilos». Biology Letters 4 (4): 415-418. ISSN 1744-9561. PMC 2610139. PMID 18426746. doi:10.1098/rsbl.2008.0138.
- ↑ Mitchell, G; Lust, A (23 de agosto de 2008). «La cola carótida y el éxito de los artiodáctilos». Biology Letters 4 (4): 415-418. ISSN 1744-9561. PMC 2610139. PMID 18426746. doi:10.1098/rsbl.2008.0138.
- ↑ Owen-Smith, Norman (1984). The Encyclopedia of Mammals. Nueva York : Facts on File, Macdonald, D. ISBN 0-87196-871-1.
- ↑ a b Hooker, J.J. 2005. Perissodactyla; in K. D. Rose & J. D. Archibald (eds.), pp. 199-214: The Rise of Placental Mammals, Origins and Relationships of the Major Extant Clades. The Johns Hopkins University Press, Baltimore. ISBN 0-8018-8022-X
- ↑ Savage, R.J.G. & Long, M.R. 1986. Mammal Evolution: an illustrated guide. Facts on File, New York. ISBN 0-8160-1194-X
- ↑ a b McKenna, M. C. & Bell, S. K. 1997. Classification of Mammals Above the Species Level. Columbia University Press, New York. ISBN 0-231-11013-8
- ↑ Mikko's Phylogeny Archive - Perissodactyla
- ↑ J. J. Hooker and D. Dashzeveg: The origin of chalicotheres (Perissodactyla, Mammalia). en: Palaeontology: Vol. 47 Part 6, 2004, S. 1363–1386 (PDF-Version)
Bibliografía
editar- Martin S. Fischer: Mesaxonia (Perissodactyla), Unpaarhufer. In: Wilfried Westheide, Reinhard Rieger (Hrsg.): Spezielle Zoologie. Teil 2: Wirbel- oder Schädeltiere. Spektrum Akademischer Verlag, Heidelberg und Berlin 2004, S. 646–655, ISBN 3-8274-0307-3.
- Thomas S. Kemp: The Origin & Evolution of Mammals. Oxford University Press, Oxford 2005. ISBN 0-19-850761-5.
- A. H. Müller: Lehrbuch der Paläozoologie, Band III: Vertebraten, Teil 3: Mammalia. 2. Auflage. Gustav Fischer Verlag, Jena und Stuttgart 1989. ISBN 3-334-00223-3.
- Don E. Wilson, DeeAnn M. Reeder (Hrsg.): Mammal Species of the World. 3. Ausgabe. The Johns Hopkins University Press, Baltimore 2005, ISBN 0-8018-8221-4.
Enlaces externos
editar Wikispecies tiene un artículo sobre Perissodactyla.
Wikispecies tiene un artículo sobre Perissodactyla. Wikimedia Commons alberga una categoría multimedia sobre Perissodactyla.
Wikimedia Commons alberga una categoría multimedia sobre Perissodactyla.
 Datos: Q25374
Datos: Q25374- Multimedia: Perissodactyla / Q25374
- Especies: Perissodactyla